Undergraduate Project (1994/5)
Alexandra Freeman

Undergraduate Project (1994/5) |
Alexandra Freeman |
|
The morphology and behaviour of gorgonopsids, and a new use for computers in palaeontology
| Aims | |
| General Introduction | |
| Material and Methods | |
| Identification | |
| General Description | |
| Reconstruction |
| Introduction to Reconstruction | |
| Thermoregulatory Status | |
| Olfaction and Other Senses | |
| Locomotion | |
| Reproduction | |
| Conclusions |
| Computer Imaging |
| Introduction to Computer Imaging | |
| Materials and Methods | |
| Results | |
| Conclusions |
| Summary | |
| Acknowledgements | |
| References |
| Appendix 1 - The FORTRAN Program | |
| Appendix 2 - Abbreviations |
When starting this project my objective was to prepare and identify a gorgonopsid skull, and to reconstruct as much as possible of the animal's morphology and behaviour in life without wandering too far into the realms of unsubstantiated speculation. Whilst attempting to do this I came across the interesting problem of how to represent a three-dimensional specimen accurately in two dimensions. So I developed a secondary aim - to produce a method of displaying a three-dimensional specimen as two-dimensional on a computer, allowing it to be viewed from any angle just as the real specimen might be.
This project therefore has two parts:
• Firstly, the identification (as far as possible) of the prepared gorgonopsid specimen, and a general description of the characters which allowed its identification, followed by an evaluation of the evidence available on which to base a reconstruction of the animal's habits in life.
• Secondly, the development of a new method of representing palaeontological specimens which could prove invaluable in making specimens more widely available for study.
Deposits from around the world show a gradual evolution from typical reptilian forms, through a series of 'mammal-like reptiles' or synapsids, to true mammals. This unique example of one class evolving from another has been well preserved and well studied. The synapsids can be divided roughly into three stages. Firstly the pelycosaurs, originating in the Upper Carboniferous, which were very reptilian, and showed the development of a temporal fenestra, providing a firmer basis for strong jaw muscle attachment. They developed both herbivorous and carnivorous habits, and showed a typical reptilian sprawling gait.
In the Late Permian, a new series of synapsids appeared, and these are preserved in large numbers in the African Karroo deposits. These animals are known as therapsids, and are characterised by the presence of a reflected lamina on the angular bone of the lower jaw, an enlargement of the temporal fenestra, and the differentiation of the teeth into incisors and postcanines separated by a single large canine in each jaw. In contrast to the tropical swampy environment in which the pelycosaurs arose, the therapsids are associated with more arid conditions with seasonal rainfall. Dominant among the early therapsids were the large dinocephalians, which adapted to form both herbivores and carnivores. In the upper Late Permian, these were completely replaced by the carnivorous gorgonopsids and therocephalians, and the highly successful herbivorous dicynodonts. In the last part of the Late Permian, the cynodonts appeared, probably from a common ancestor shared with the therocephalians, showing many mammalian features such as multicusped postcanine teeth. The cynodonts persisted through the Triassic, and eventually gave rise to the true mammals which formed the dominant fauna after the demise of the dinosaurs.
The gorgonopsids are thought to be related to certain poorly known primitive therapsid carnivores such as Phthinosaurus, Phthinosuchus, Biarmosuchus, and Eotitanosuchus. Possibly the most primitive animals which can be directly linked to the gorgonopsids are the ictidorhinids from the Karroo deposits, which have many primitive features resembling Biarmosuchus and Eotitanosuchus, but are not yet known in detail. They are classified as the Infra-order Ictidorhinae, as opposed to the Infra-order Gorgonopsia, which contains the family Burnetiidae, known only from two skulls bearing bony protruberances but with dental and palatal similarities to gorgonopsids, and the conservative family Gorgonopsidae. The characteristic gorgonopsid features are generally adaptations to catching and feeding on large prey. They include the development of even larger 'sabre' canines and interdigitating incisors, a short, deep snout to accommodate the teeth, and enlarged temporal fenestra in order to allow the further strengthening of the jaw musculature. Two other characteristic features are the presence of a preparietal bone, and the meeting of the palatine bones at the midline. The Gorgonopsidae are often divided into two sub-families, the Rubidginae, which had large, broad skulls, and the Gorgonopsinae, which comprised most of the gorgonopsid genera. These are usually distinguished only by characters such as the number of postcanines, and the various proportions of the skull (Kemp, 1982; Hopson and Barghusen, 1986).
| Infra-order Ictidorhinae |
| Infra-order Gorgonopsia |
| Family Burnetiidae | |
| Family Gorgonopsidae |
| Subfamily Rubidginae | |
| Subfamily Gorgonopsinae |
At the end of the Late Permian, the gorgonopsids became abruptly extinct leaving no descendants, and were replaced by the cynodonts which eventually gave rise to true mammals. The gorgonopsids' place in the middle of the series from reptiles to mammals makes them particularly interesting. By studying their physiology and anatomy we may understand more fully how the transition from reptiles to mammals was achieved, and further how the process of evolution acts to produce such radical changes.
The material consists of a large gorgonopsid skull, slightly flattened dorso-ventrally, and sheared from left to right, together with a quadrate bone, a near-complete right femur, a partial left humerus, a left tibia, at least three vertebrae, and other parts of the post-cranial skeleton. The material is from the Madumabisa Mudstone region of the Karroo System of mid-Luangwa Valley, Northern Zambia (Kemp, 1975). The collection was made by Dr. T. S. Kemp in 1974, and is housed in the University Museum, Oxford. This specimen is number T.S.K. 91. After a comparison of the sizes of the skull and the postcranial fragments, it appears that the skull is not from the same animal as the associated skeleton.
The skull lacks a lower jaw, although a quadrate is present. The teeth are missing, although most of the sockets are visible. The vomer and the ventral keel of the palate are missing, and the palate can be removed from the rest of the skull. The squamosals are damaged on both sides, as are the sub-orbital bones on the right hand side. The specimen was preserved largely free of matrix, but that remaining was removed by careful use of abrasive apparatus, and the specimen assembled using 'Vinalak' (polybutyl methacrylate).
The large broad skull and broad snout comprising approximately half the skull length, is characteristic of the subfamily Gorgonopsinae of the Gorgonopsia (Sigogneau-Russell, 1989). The large size of the skull, its proportions and extensive ornamentation suggest that it is at least closely related to Leontocephalus intactus (Kemp). As the genera Leontocephalus and Leontosaurus are both somewhat disputed, and the type specimen of Leontocephalus intactus is not available for comparison, this identification can only be tentative.
The skull has suffered slight crushing dorso-ventrally, and has undergone a shearing force from left to right. It is large and heavily built, and displays much ornamentation until just behind the orbits. The upper surfaces of the nasals are covered in longitudinal pits, which decrease in prominence from the anterior to the posterior and cease at the anterior margins of the orbits. The margins of the orbits are very heavily rugose, but the frontals, prefrontals and postfrontals are less so. The lateral surfaces of the maxillae are slightly rugose, but the dorsal margins are smooth. The parietals and other bones are also smooth. The pineal is surrounded by a raised boss, and just anterior to it is a similar, slightly raised area which extends a few centimetres in a low, slight crest. This area, however, has been damaged by crushing, and the structures are not clear. See Figs. 1 and 2.
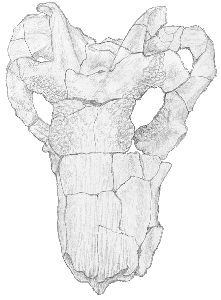 | 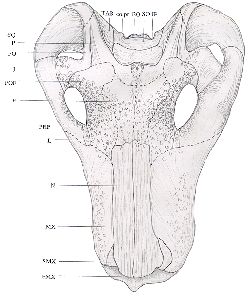 |
| Fig. 1 Dorsal view of skull (x 1/2) |
| Fig. 2 Reconstructed dorsal view of skull (x 1/2) (Dashed lines represent possible routes of sutures not visible) |
In ventral view the distortion of the skull is obvious, as the sagittal line of the skull has been displaced to the right in the dorsal region, whilst the ventral structures remain more regular. The premaxilla is partially damaged, and only the remains of three incisors remain, on the right side. A small depression on the left side may be the remains of an incisor socket. Five incisors were probably originally present on each side. The right maxilla bears a large canine socket, the remains of two small post-canines, and just anterior to them the possible remains of a third post-canine socket. The corresponding area on the left has been damaged. The vomer is entirely missing. With the palate removed, the turbinal ridges on the internal surfaces of the nasal and frontal bones can be seen. See Figs. 3 and 4.
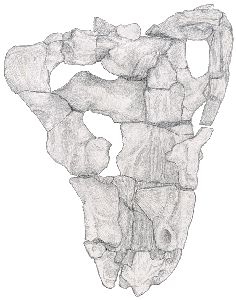 | 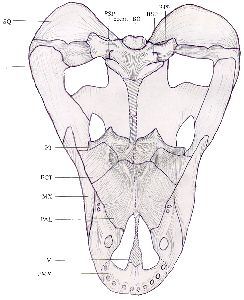 |
| Fig. 3 Ventral view of skull with palate removed (x 1/2) |
| Fig. 4 Reconstructed ventral view of skull with palate in place (x 1/2)(Hatched areas represent reconstruction of missing bone) |
With the palate in place, the ectopterygoid can be seen to be large and to form an important part of the junction between the maxilla and the palate. The maxilla meets the palatine and ectopterygoid without leaving any space, and there is a process of the maxilla overlying the jugal. The palatine bosses are quite large, and rugose. The pterygoid bosses are similar, although slightly smaller. There are no teeth visible on either pair. The ventral keel behind the palate is missing from the pterygoidal flanges to the parasphenoidal tubera. These tubera are large and rugose, but the basisphenoidal tubera are flat and relatively smooth. The paroccipital processes are poorly preserved, but appear to be flat and wide. The opening of the parietal foramen is obscured by the parasphenoid, the anterior portion of which is missing. The ventral region of the left side is flattened dorso-ventrally from the anterior margin of the orbit until the posterior margin of the skull. See Figs. 4 and 5.
 |
| Fig. 5 | (a) Ventral view of palate (x 1/2) |
| (b) Anterior view of palate (x 1/2) |
In lateral view, from the right side, the dorsal margin of the skull is almost straight, as the crushing has not affected this edge. There appears to be a slight step between the last incisor and the canine on the ventral margin, but this region is damaged on both sides, and the suture between the maxilla and premaxilla is obscured. The jugal overlaps the squamosal extensively, and a flange of the maxilla overlies the jugal posteriorly and very slightly internally. The temporal fenestrae are shallow and broad. The dorsal edge of each is bounded by the post-orbital, which is perfectly smooth, and there is a longitudinal furrow along which the suture of the parietal and post-orbital lies. See Fig. 6.
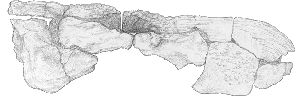 |
In occipital view, the crushing and left to right shearing of the skull is obvious, as is its generally flattened character. Due to the crushing, the region is not well preserved, but seems to correspond to Kemp's (1969) description of Leontocephalus intactus. See Fig. 7.
 |
(After Sigogneau, 1970). Due to the distortion of the skull, many of the measurements are likely to be inaccurate.
| lg. mx. | = 370mm | (maximum length from end premaxilla to end squamosal) |
| la. mx. | = 320mm | (maximum width between zygomatic arches) |
| lg. mu. | = 170mm | (maximum length snout from premaxilla to before orbits) |
| la. mu. | = 150mm | (width snout 2/3rds from anterior end) |
| ht. mu. | = 130mm | (height snout 2/3rds from anterior end) |
| io. | = 100mm | (minimum inter-orbital width) |
| it. | = 120mm | (minimum inter-temporal width) |
| ob. | = 70mm | (length of orbits) |
| pin. | = 280mm | (length from end premaxilla to pineal hole) |
| pin. cr. | = 80mm? | (distance pineal hole to nuchal crest) |
| a. tv. | = 190mm | (distance end premaxilla to nearest edge of peak convexity transverse pterygoid apophyses) |
| pob. | = 45mm | (minimum width post-orbital bar) |
| s. ob. | = 35mm | (minimum height sub-orbital ridge) |
| zyg. | = 25mm | (minimum height zygomatic arch) |
| de. | = --- | (height dentary behind last post-canine) |
| I5.C. | = 20mm | (length diastema fifth incisor to canine) |
| C. | = 20mm? | (maximum diameter functional canine) |
| C-Pc. | = 35mm | (length diastema canine to first post-canine) |
| Pc. | = --- | (length post-canine series) |
| f.t. | = 95mm | (maximum length temporal fossa) |
Faced with a fossil skull and a few fragments of the post-cranial skeleton, it seemed a challenge to try to reconstruct as much as possible about the morphology and behaviour of the gorgonopsids, and in particular to attempt to form an image of how the specimen described might have looked and acted when it was alive. Various aspects of gorgonopsid anatomy have been studied in detail by others, and this section mainly acts as an evaluation of their work based on my own observations and thoughts after an examination of the specimen available to me. I have not repeated in detail the findings of other authors, but instead have tried to restrict my conclusions to those which have a direct bearing on the appearance or behaviour of the animal.
Mammals are endothermic - maintaining a high basal metabolic rate by the production of internal heat. However, they evolved from ectothermic reptiles, and the mammal-like reptiles must therefore show a change in thermoregulatory status through time. It is very difficult to infer physiological characters from fossilised structures, and the fact that modern endotherms and ectotherms are so different from each other in many respects makes the association of certain characters with endothermy a difficult task. Bennett and Ruben (1986) have evaluated the evidence often cited for the presence of endothermy, and I have applied their conclusions to the Gorgonopsia in an attempt to determine their thermoregulatory status.
Highly vascularized, metabolically active bone is suggestive of endothermy, but the relationship is not direct, and therefore the interpretation is controversial. Bone structure can be divided broadly into two types - lamellar-zonal, where the bone is poorly vascularized, has a layered appearance, and there are few primary osteons; and fibro-lamellar, where many primary osteons lay down most of the matrix, which has a woven appearance, is well vascularized, and often forms extensive Haversian systems. Lamellar-zonal bone is usually associated with ectothermy, and fibro-lamellar bone with endothermy, but the categories are not distinct, and many factors (such as habitat and size of the individual) can affect the structure of the bone (Bennett and Ruben, 1986).
Thin longitudinal and transverse sections had already been taken through the femur of a large, unidentified gorgonopsid from the same collection as the specimen described, and the histology of this unidentified specimen (T.S.K. 53) was studied (See Fig. 8).
 |
| Fig. 8 | (a) Transverse section through gorgonopsid femur shaft (x 17) | (b) Longitudinal section through gorgonopsid femur shaft (x 9) |
As the specimen was large, and terrestrial, the problems arising from small, aquatic, or aerial species, which often have specialised bone structures not corresponding to generalised statements, do not affect the conclusions. The structure corresponded closely with that observed by RicqlŽs (1969). The outer layer, the cortex, is made of dense compact bone, and is relatively thin. Longitudinal vascular channels pass through this, arranged in four or so concentric circles. These form primary osteons, but secondary osteons are rare. The bone thus cannot be assigned to either category, but the absence of extensive Haversian canals, and the presence of apparent growth rings in the pattern of growth, suggests that the Gorgonopsia were in fact ectothermic, and that their growth was affected by seasonality.
A fully upright stance is often cited as evidence for endothermy. Some authors (e.g. Heath, 1968) argue that there is a causal correlation between the two, due to the increased muscle use, and thus heat production. However, endothermic heat is primarily produced by internal organs rather than skeletal muscle, and thus a causal connection must be at least unlikely. Modern animals also do not show a perfect correlation. Crocodiles, for example, show a semi-erect stance, and yet are fully ectothermic. The stance of the gorgonopsids is itself disputed, and this will be discussed later.
Hair is strongly linked with endothermy, a full pelage being primarily beneficial as insulation to an endotherm. Foramina on the snouts of gorgonopsids (among other therapsids) have been interpreted as passages for the supply of blood and nerves to facial vibrissae. However, identical foramina are found in the skulls of living ectotherms, with no vibrissae and scaly skin. Only one piece of fossilised skin has been found, from the dinocephalian Estemmenosuchus, which shows a smooth, undifferentiated surface bearing no scales or hair follicles. The skin was, however, well supplied with glands, and was probably soft and pliable (Chudinov, 1970, cited in Bennett and Ruben, 1986). The skin may thus have resembled that of a rhinoceros. This suggests that scales may already have been lost in the Gorgonopsia, which diverged from the mammalian line later than Estemmenosuchus. The presence of many skin glands also opens the possibility of communication by scent, as the presence of oily substances on the surface of the skin, to keep it pliable, is likely to have given the animals a distinctive smell, which could be exploited by predators and the animals themselves for intra-specific signalling. This will be discussed later. The lack of direct evidence for hair or scales, however, gives no clue as to thermoregulatory status in the Gorgonopsia.
The presence of a causal connection between high metabolic rate and a high degree of encephalisation is greatly disputed, and estimations of the brain size of Mesozoic mammals places them between those of modern reptiles and modern mammals (and close to those estimated for therapsids). Thus, there was no large jump in brain size accompanying endothermy, and degree of encephalisation cannot be used as evidence for thermoregulatory status (Ulinski, 1986).
5) Presence of a diaphragm
A muscular diaphragm can increase the efficiency of lung ventilation, and thus allow a higher metabolic rate in animals which possess one. Its presence can only be inferred from the reduction in lumbar ribs. This reduction could, however, also be associated with increased agility in locomotion, and the presence of a diaphragm-like structure (such as that found in crocodiles) does not necessarily indicate endothermy.
Gorgonopsids are considered not to have reduced lumbar ribs, and as they were running animals, and would not have retained these ribs for support purposes if they had required a diaphragm (as in some burrowing animals), we can tentatively conclude that gorgonopsids did not possess a diaphragm, and that this possibly precludes them from having a high basal metabolic rate.
It has been argued that a bony palate, separating the buccal and nasal cavities, allows breathing during feeding. This represents the need for continuous ventilation in endotherms. However, some ectotherms (e.g. crocodiles) possess a bony palate, and some endotherms (birds) do not. The separation of the air and food passages appears to be effective if the palate is fleshy or cartilaginous, and the ossification of it serves only to allow manipulation of the food against it by the tongue.
The gorgonopsids did not possess a bony palate, although it may have been cartilaginous. Although this gives no evidence as to their thermoregulatory status, it does suggest that the food tended to be swallowed without mastication. This is supported by the lack of grinding post-canine teeth. This in itself is indicative of a primarily reptilian metabolism, as endotherms require a greater efficiency in energy extraction from their food, which is achieved partly by mastication of the food in the mouth allowing enzymes to act over a greater surface area.
Modern mammals possess extensive nasal turbinals, whose moist epithelia warm inhaled air and reclaim water from exhaled air. These turbinals are only present in living endotherms, and the conditioning of air would be unnecessary in an ectotherm; thus Bennett and Ruben (1986) suggest that the presence of nasal turbinals is the best evidence available for endothermy.
Evidence for nasal turbinals is present in the specimen described, and Kemp (1969) further describes a gorgonopsid skull in which evidence of cartilaginous turbinals is preserved in a mineralised form. Thus, either these gorgonopsids were endothermic, or the nasal turbinals were present for some other reason, such as the detection of scents in the air. This will be discussed later.
The parietal foramen provides an aperture for the parietal eye which, together with the pineal body, forms the epiphyseal complex. Sphenodon and some lizards show this complex in its complete form, but in most tetrapod lineages the external connection has been lost, and only the pineal body retained. The function of the epiphyseal complex is not fully understood, but it appears to be associated with behavioural thermoregulation in ectotherms (Roth, Roth, and Hotton, 1986). As its function is not understood, the retention or loss of a parietal foramen cannot definitely be ascribed to any thermoregulatory cause.
The specimen described shows a parietal foramen, in common with other gorgonopsids, and this may be very tentative evidence for the presence of ectothermy in this group.
As endotherms have a much higher energy-flow, and thus require more energy-rich food, the predator-prey ratio for endotherms is generally much lower than that for ectotherms. However, there can be much overlap in this respect. The difficulties involved in estimating the relative biomasses of predators and prey from the fossil record make this a very unreliable source of evidence. However, carnivores appear to make up less than 5% of the fossil remains - possibly evidence for endothermy.
Bakker (1975, 1980) has argued that large ectotherms could not have survived in Southern Gondwanaland (now forming the South African Karroo deposits), as this area was at high latitudes in the Late Permian. However, evidence from tree-rings and other sources suggests that by the Early Permian ice sheets had retreated from this area, and the climate was warm and showed wet-dry seasonality. The presence of large animals which were definitely ectothermic (such as the early thecodonts) in the fauna together with the therapsids demonstrates that the climate did not restrict the evolution of large ectotherms. Thus, biogeography can give no clue as to the thermoregulatory status of the Karroo deposit fauna.
The conclusions which can be drawn from this evidence are still somewhat tentative, as evidence from different characters is contradictory, and it is impossible to weight the evidence satisfactorily. The evidence can be summarised as follows:
| Evidence for endothermy | Evidence for ectothermy |
| Nasal turbinals - strong? | Bone histology - quite strong |
| Predator-prey ratio - weak | Lack of diaphragm - quite weak |
| Lack of mastication - quite weak | |
| Parietal foramen - weak |
The number of different sources of evidence suggesting that the Gorgonopsia were ectothermic may outweigh the strength of the evidence from the presence of nasal turbinals which suggests that they were endothermic. If this is so, then the nasal turbinals must have had some function other than to condition the air entering and leaving the lungs. I suggest that this is the case, and that the Gorgonopsia were ectothermic.
If the Gorgonopsia were indeed ectothermic, then the presence of bony ridges implying large nasal turbinals requires an explanation. Modern reptiles do not possess nasal turbinals under any environmental conditions, and thus their use to filter out dust or water vapour in the atmosphere seems unlikely for the Gorgonopsia, especially as the climate of Southern Gondwanaland at that time was thought to be equable and not particularly dry or dusty. It has been suggested that the large nasal turbinals were instead used to detect scents in the air (Graves and Duvall, 1983). Gorgonopsids were carnivores, and probably fed mainly on the numerous herbivorous dicynodonts of that time. The fragment of fossilised skin from Estemmenosuchus mentioned previously appeared to show many sebaceous glands, which indicates that these animals were likely to give off distinctive scents. If that was indeed the case, the Gorgonopsia would greatly benefit from being able to detect their prey by scent, and I suggest that the nasal turbinals were used for this purpose. Whether or not the animals used pheromones for intraspecific communication is a different matter. There is no evidence to indicate the nature of the gorgonopsid skin, but it is possible that it resembled that of the dicynodonts. If so, intra-specific scent communication is likely to have evolved, as the mechanism for it would already have been present for use in detecting prey.
There is physiological evidence to indicate that the nasal turbinals did not in fact affect the respiratory air (Kemp, 1969). Instead, the nasal capsule was divided into two chambers by a pair of processes of the palatine bones. The anterior chamber is relatively small, and the air entered it via the external nares, being deflected ventrally through internal choanae and back down the respiratory passage. The posterior capsule, by contrast, is very large, and contains the nasal turbinals. It is supplied with air by the septomaxillary foramina which passes through the anterior capsule, possibly being channelled by cartilaginous structures, which would also rechannel the air back into the anterior capsule as more air was inspired. If this interpretation is correct, then the large posterior cavity containing the cartilaginous turbinals must indeed have been used for olfaction, and hence the Gorgonopsia would have had a sensitive scent reception mechanism.
The vomeronasal or Jacobson's organ is another structure for odour reception, and is thought to have been present in therapsids as it is a primitive tetrapod structure retained in some mammals (Duvall, 1986). It is used particularly in the detection of intraspecific social and sexual pheromones. Its presence in cynodonts can be inferred by the presence of incisive foramina in the secondary palate (Duvall, 1986; Duvall, King, and Graves, 1983). In the Gorgonopsia, which lacked a bony palate, there is no evidence for a vomeronasal organ, but it can be assumed to have been present. If this is the case, the enlarged olfactory area of the nasal cavity was probably used in interspecific scent recognition and prey detection. In the absence of evidence relating to the size of the vomeronasal organ in the Gorgonopsia, it is impossible to postulate the importance of intra-specific scent communication in these animals. If they were indeed ectothermic, then it is possible that they used pheromones to a similar extent to modern day terrestrial reptiles.
The importance of olfaction has already been discussed. The orbits are dorsally placed, but the head was probably carried at an angle, with the snout pointing downwards as in dogs. Thus the snout would not obscure vision, and the eyes would be forward pointing. Vision and coordination must have been important for gorgonopsids as fast running predators (see 'Locomotion'). The importance of hearing is much less certain, although the stapes is lightly built, and rests in a recess in the quadrate bone without firm attachment. This seems to suggest that it did have some use as a sense structure, although it would be relatively insensitive (Carroll, 1986), and could probably only detect low frequency sounds. Thus the gorgonopsids probably used sight and smell to detect prey, and hearing only to a slight extent such as the movement of herds of dicynodonts. Their reproductive signalling probably involved pheromones detected by the vomeronasal organ, as in modern reptiles, and possibly visual courtship.
The gorgonopsids in general are lightly-built with long, slender limbs. This would seem to be an adaptation for fast running, and prey capture. There has, however, been some debate as to their gait. Reptiles tend to employ a 'sprawling' gait, where the humeri and femora are held horizontally. This involves a degree of lateral undulation of the body, and is generally a relatively slow means of locomotion. Mammals, by contrast, demonstrate an 'upright' stance, where the humeri and femora are held in a parasagittal plane. This is a faster means of locomotion, involving some dorso-ventral flexing of the spine. The two types of locomotion have been shown to require similar amounts of energy, and thus are not directly correlated with metabolic status.
The mammal-like reptiles are likely to show a trend for increasingly upright stance, but this could take two forms. Either the animals could have moved through a series of semi-erect stances until the mammalian condition was reached, or they could have employed dual gaits such as is seen in modern crocodilians. Crocodilians show a sprawling gait for slow locomotion, and a semi-erect stance for faster movement. It is possible that some of the intermediate therapsids employed dual gaits, and gradually the upright stance became favoured for locomotion in general (see Sues, 1986).
These differing forms of locomotion use different muscles, and thus have an effect on the shape of the limb bones. Comparative studies of the gorgonopsid limb bones have been carried out in order to predict the locomotory methods of these animals (e.g. Colbert, 1948; Gale, 1985). The femoral head of gorgonopsids is turned inwards, indicating that the femur was not held horizontally. However, there is no femoral neck, which would indicate a fully upright stance, so the osteological evidence favours a semi-erect gait. Examination of the femur found in association with the skull previously described shows the large trochanter major, indicating the presence of a well-developed ilio-femoralis muscle, which is used in the sprawling gait as a rotator about the long axis of the bone, and in an upright gait as a retractor muscle (See Fig. 9). The twist in the shaft of the bone is very similar to that seen in crocodilian femora, and allows the ilio-femoralis muscle to carry out both these functions without damage, thus facilitating adoption of both gaits at different times. The internal trochanter is long and well-developed, but does not form a prominent ridge along the ventral surface. This indicates that the puboishiofemoralis muscle (PIFE), which acts as an adductor in a sprawling gait, was strong, but a semi-erect gait could be adopted as well.
 |
Gale (1985) analysed the micro architecture of a gorgonopsid femur, and concluded that both a sprawling and semi-erect gait could have been used. The femora also show a superficial resemblance to crocodilian femora, although the relative sizes of the trochanters are different. The ankle joint also shows some specialisation reminiscent of the crocodilians in being more generally mobile, allowing different gaits to be employed, with the foot being plantigrade. In addition, the lack of intercentra between the vertebrae (apart from the cervical vertebrae) indicates that lateral undulation, and thus the use of the sprawling gait, was reduced (See Fig. 10). The humeri are similar to the ancestral pelycosaurian condition, and would have been held in a similar, horizontal manner. Thus the gorgonopsids could have employed either a permanent semi-erect gait, or showed both sprawling and semi-erect locomotion from the hind limbs, providing a faster prey-catching force when necessary.
 |
| Fig. 8 | (a) Lateral view of two vertebrae found in association with the skull | (b) Anterior view of one of the vertebrae found in association with the skull |
The skeleton is generally lighter, and more adapted for manoeuvrability during fast locomotion when compared to pelycosaurs. The gorgonopsids can thus be seen as likely to have been fast-running predators, although they were probably ectothermic, and as such lacked the stamina of mammalian predators. Their enlarged canines and wide gape would allow them to deliver a disabling bite to prey of any size. The pterygoideus muscle used for jaw-closing at wide gapes was particularly large (Kemp, 1969) indicating that this was indeed the case. The movements of the jaw have been explored in depth (Kemp, 1969), showing that propaliny of the lower jaw led to interdigitating of the incisors, which would have sliced swallowable chunks of meat off the prey. This action was also undertaken by the powerful pterygoideus muscle.
If gorgonopsids were indeed ectothermic, they would be likely to have shown typical reptilian reproduction, with none of the mammalian characters such as mammary glands or social bonding (Guillette and Hotton, 1986). As they were active predators, it seems likely that the females laid eggs, which they then abandoned (at least for a while), as the cost of ovovivipary or vivipary would be too high. The female would need to hunt quite frequently, and carrying a brood would put unnecessary strain on her ability to catch prey. The eggs may have been buried for safety, as seen in many modern reptiles, in which case the mother may have been required to unearth the hatching eggs, and protect the young for a short period, as do modern crocodilians.
By comparing the sizes of the skull described (T.S.K. 91) and the associated post-cranial material, the two can be seen to be from animals of quite different sizes. However, from the conclusions drawn about the behaviour of gorgonopsids and the detailed reconstruction of Lycaenops ornatus (Broom) by Colbert (1948), a possible image of the specimen T.S.K. 91 in life can be obtained, assuming that the proportions of the head and the rest of the body is similar to that in Lycaenops (see Fig. 11).
The specimen to which the skull belongs would probably stand about 0.6m high and would be about 1.75m long when in the pose illustrated (see Fig. 11), whilst the specimen to which the post-cranial remains belong would probably have stood about 0.33m high, and been 1m long (by comparison with the proportions of Lycaenops), and would have had a semi-erect gait when moving fast. In general, it would probably look rather crocodilian in locomotion. The tail would also be thick, as in crocodilians, due to the ilio-ischio-caudalis muscle, but would not be as long and exaggerated as it is in crocodilians, as it would not be used for swimming. The head is proportionately very large, and would probably be carried rather like that of a dog. The skin was probably hairless, and possibly smooth, rather like a rhinocerous (see Fig. 11b).
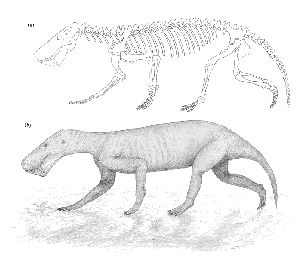 |
| Fig. 11 | (a) Reconstructed skeleton of T.S.K. 91 based on post-cranial skeleton of Lycaenops (Colbert 1948) and the skull of Leontocephalus (Kemp, 1969) | (b) Possible image of the specimen in life |
Gorgonopsids were likely to be solitary hunters, as they would not have the cranial capacity or means of bonding to facilitate complex group interactions. They probably detected herbivorous dicynodonts by scent and vision, and made a short sprint to catch them, disabling their prey with a powerful bite with the enlarged canines. The meat could then be ripped off using the jaw propaliny, and swallowed in chunks.
Due to the seasonal wet-dry climate of Southern Gondwanaland at the time, the gorgonopsids were likely to have been annual breeders, locating mates by vision and the detection of pheromones using the vomeronasal organ. The females probably laid eggs, possibly buried for protection. There is also the possibility of a degree of parental protection for the young, especially if the mother returned to the nest site to unearth hatchlings as crocodilians do.
Specimens that are distorted in some way are usually reconstructed simply by the use of imagination by the artist, and my original idea was that this could be more accurately done by correction on a computer. My initial attempts in two dimensions proved unsuccessful, as the complex forces involved could not be resolved into correcting a two dimensional portrayal (a drawing scanned into an Apple Macintosh computer and manipulated using the 'shear' command in SuperPaint). As I investigated the possibility of three dimensional representation, it became clear that the conversion of a specimen into a series of (x, y, z) coordinates, and their subsequent portrayal on a grid posed interesting problems which had previously not been tackled.
I first contacted the Engineering Department, Oxford University, in order to determine whether or not an instrument existed which could digitise an object in three dimensions, and eventually found that such an instrument had just been acquired for use in the Nuffield Orthopaedic Centre, Headington, Oxford. The digitiser had just been calibrated, but had not yet been used, as there was no computer software available to plot the coordinates once they had been obtained. I had found an Apple Macintosh program (VISION-3D, written by Paul Bourke) which appeared to be able to plot coordinates in a three-dimensional array, and join the nearest neighbours (using a mathematical technique known as Delauney triangulation). As the Orthopaedic Centre were very interested in the possibility of using the digitiser to produce coordinates which could then be graphically manipulated (in the production of prosthetic limbs), they kindly allowed me to use the instrument.
The coordinates were manipulated into a form which could be read by VISION-3D, and a Delauney triangulation attempted. However, even a small fraction of the points proved to be too many for the capabilities of the program, and I contacted the Oxford University Computing Services for help. A software system known as AVS exists which can carry out many imaging techniques. However the mathematics of the problem of joining the array of points proved insuperable, as there is no unique way of joining a 'random' array. Delauney triangulation would be unsuitable. So, a FORTRAN teaching program was altered in order to simply plot the points in a three-dimensional grid. By further alterations to the program, the size of the dots was altered, and the array sorted into layers depending on their distance from the viewpoint, which could be plotted in different colours to give a perception of depth in the object. By plotting the dots relatively large, they could be made to merge into each other, giving the illusion of a solid object. In addition, the layers can be plotted singly (in effect, looking at a cross-section through the specimen), and the viewpoint moved to look at the specimen from any angle or distance.
Thus, to my knowledge for the first time, a palaeontological specimen can be portrayed accurately as a three-dimensional object on a computer screen, and then manipulated in a way that the original specimen could not (such as slicing). This technique could prove very valuable in palaeontology, where access to specimens is limited, and further refinements to the programming could allow more accurate reconstruction of distorted specimens. The possibility of cross-sections could be particularly valuable in the analysis of mechanical stress on bones, and thus the reconstruction of muscle action and behaviour in extinct animals.
The prepared specimen (see previous description) was digitised using an ISOTRAK-II position tracker manufactured by POLHEMUS (Vermont, U.S.A.). This apparatus produces a magnetic field around a surface made entirely from non-magnetic substances onto which the specimen is placed. A small, hand-held electronic pen with a metal tip is then placed on the surface of the specimen, and a button pressed to take a reading. The coordinates of the pen in 3D space are recorded relative to the artificial magnetic field, and displayed on an associated IBM computer screen. The coordinates are automatically stored on the computer's hard disc.
The specimen was placed in the magnetic field in the positive z-range (running from the posterior to the anterior), positive y-range (running from the ventral to the dorsal surface), and with the x-axis running down the midline. The most distal point, at the tip of the snout, was chosen as a reference point, and marked in pencil on the specimen. This point was taken as the first reading, and subsequent readings were taken moving from the anterior to the posterior over the entire dorsal surface. This data was stored in two separate data sets. The specimen was then inverted, so that the ventral surface was uppermost. Care was taken to ensure that the skull remained otherwise in exactly the same position as in the previous readings. The reference point was again taken as the first, and this time also the last, reading in each data set. This allowed comparison of the first and last readings to ascertain whether or not the specimen moved position slightly during the readings. Again, as much of the ventral surface as possible was covered, and stored in two separate data sets. Finally, the palate was placed in position on the ventral surface of the specimen, and readings taken over its ventral surface stored in a fifth data set. In total, just over 20,000 readings were taken. The dorsal surface was covered with a density of around four readings per square centimeter, the ventral surface and palate at a lesser density.
The data was converted to be read by an Apple Macintosh computer, and manipulated using WORD 5 into three columns of figures (x-, y-, and z-coordinates) separated by blanks. It was imported into MINITAB in this format, with each of the five data sets consisting of three columns. The data sets were displayed alongside each other, allowing a comparison of the reference points. The reading from the first dorsal surface data set was taken as the 'true' reading, and the second dorsal data set adjusted by simple addition of the difference between the first figure in the first data set and the first figure in the corresponding column of the second data set to the whole of the column in the second data set. The z-coordinates for the ventral data sets were adjusted in the same way. The x-coordinates for the ventral data sets were multiplied by (-1) to rotate them around the x-axis. The z-coordinates were rotated around the axis containing the true reference point by multiplying them by (2r-z), where z represents the z-coordinate of the reference point of the ventral data set and r the z-coordinate of the true reference point. Extra complications arose, as some of the coordinates had negative z-values (which could not be points from the skull, which was contained entirely within the positive z-range). These readings included two of the initial reference points of the ventral data sets. Thus, the terminal reference points were used to manipulate the data. The other, negative z-value points were non-random, but seemed inexplicable, and not just rotated about the y-axis. These points were removed from the data (they totalled about two hundred), and the five data sets merged into one. The anomalous points were later discovered to be due to the digitising method employed by the position tracker.
The points were then plotted in 3D using a FORTRAN program in UNIRAS. This program sorts the data into an array, and assigns a specified colour to each point depending on its distance from the viewpoint (which is specified). The points can be plotted any size, so that they can be made to simulate a 3D model of the specimen, using the axes and colours as perspective guides. The array can be viewed from any point, and sectioned.
Although the data is rather patchy, the following four graphs demonstrate the capabilities and potential of the technique. In each graph, the red dots represent points closest to the eye, and the violet dots points furthest from the eye. The spectrum in between represents different layers of distance from the eye.
The first graph is an angled dorsal view of the skull, most clearly showing the shape and the way that points close together can give the image of a solid object. The snout is on the left, and the squamosals extend to the right.
The second graph is a lateral view, which can be compared with Fig. 6. The representation is not entirely accurate because of the difficulties involved in transforming the data, which could be overcome as is discussed later.
The third graph is a large scale dorsal view of the skull, which shows the scarcity of the data points (the first graph, however, demonstrates how even this low density of points can give the image of a solid object when viewed from a greater distance). The shape can be compared with that of Fig. 1.
The fourth graph represents a cross-section through the posterior region of the skull. The central region is the braincase, which is not externally visible.
The plots clearly show the general shape of the skull, although many areas can be seen to be poorly represented. This is probably due to some problem in the digitiser. The instrument could sometimes be rather insensitive, and may not have read some points, or some points could somehow have been lost in the memory. The view from directly above also seems to show some slippage between the data sets. This is due to a lack of reference points. For a three-dimensional structure, at least three reference points should have been taken, and I recommend that in future about five are taken from a variety of areas on the surface of the specimen.
If the specimen had been smaller, it could have been digitised much more accurately and quickly, and the representation of it would have been correspondingly more informational. The number of levels of depth represented (limited in this case by the number of available colours in the UNIRAS system) would also give a more detailed picture of the surface of the specimen. The section through this skull gives an idea of the possibilities of the technique, in giving a slice through a specimen which could not otherwise be seen.
If a small, delicate specimen were to be digitised and represented on a computer using this technique, it could easily be viewed by anybody without the risk of damage, or the need to transport more than computer data. In addition, the image of the specimen could then be manipulated on the screen in a way in which the original specimen could not. With greater refinements, large specimens could also be represented accurately, and possibly manipulated mathematically to correct for damage to the specimen (such as crushing). Thus there is the potential to make specimens more generally available to those interested, and to save a large amount of money presently spent on transport of specimens and those interested in them.
The accurate portrayal of cross-sections through specimens could be useful in the analysis of mechanical properties of bones. By studying the thicknesses of different areas of bone, the forces applied to them could be predicted and compared, thus allowing more accurate reconstructions of musculature and behaviour of extinct animals. This could be of particular use in the study of jaw mechanisms, where the shape of the skull makes the study of bone thicknesses particularly difficult.
Thus the method outlined could, with further refinements, become a very important tool in the study of palaeontological specimens, and allow a much wider appreciation of some specimens, the limited availability of which is currently a constraint on the amount of work that can be done on them.
In completing this project, I have cleaned, prepared, described and identified a gorgonopsid specimen, and reconstructed its morphology and behaviour as far as possible from the available data. I have also developed a new technique for displaying specimens, which could allow important advances in palaeontology by making specimens more widely available for study as computer representations. The technique allows sections to be taken through the representations at any angle without harm to the specimens allowing the various thicknesses of bone to be measured accurately, possibly aiding the analysis of mechanical stresses applied to various parts of the bone, and thus reconstruction of the musculature. With further modifications, the computer representations could be subjected to mathematical transformations in order to correct distortions caused during the fossilisation period, and also to demonstrate the relationships and movements between associated bones.