Alexandra Freeman
| Address: | Oxfordshire, UK |
| e-mail: |
|
| Twitter/X: | @alex_freeman |
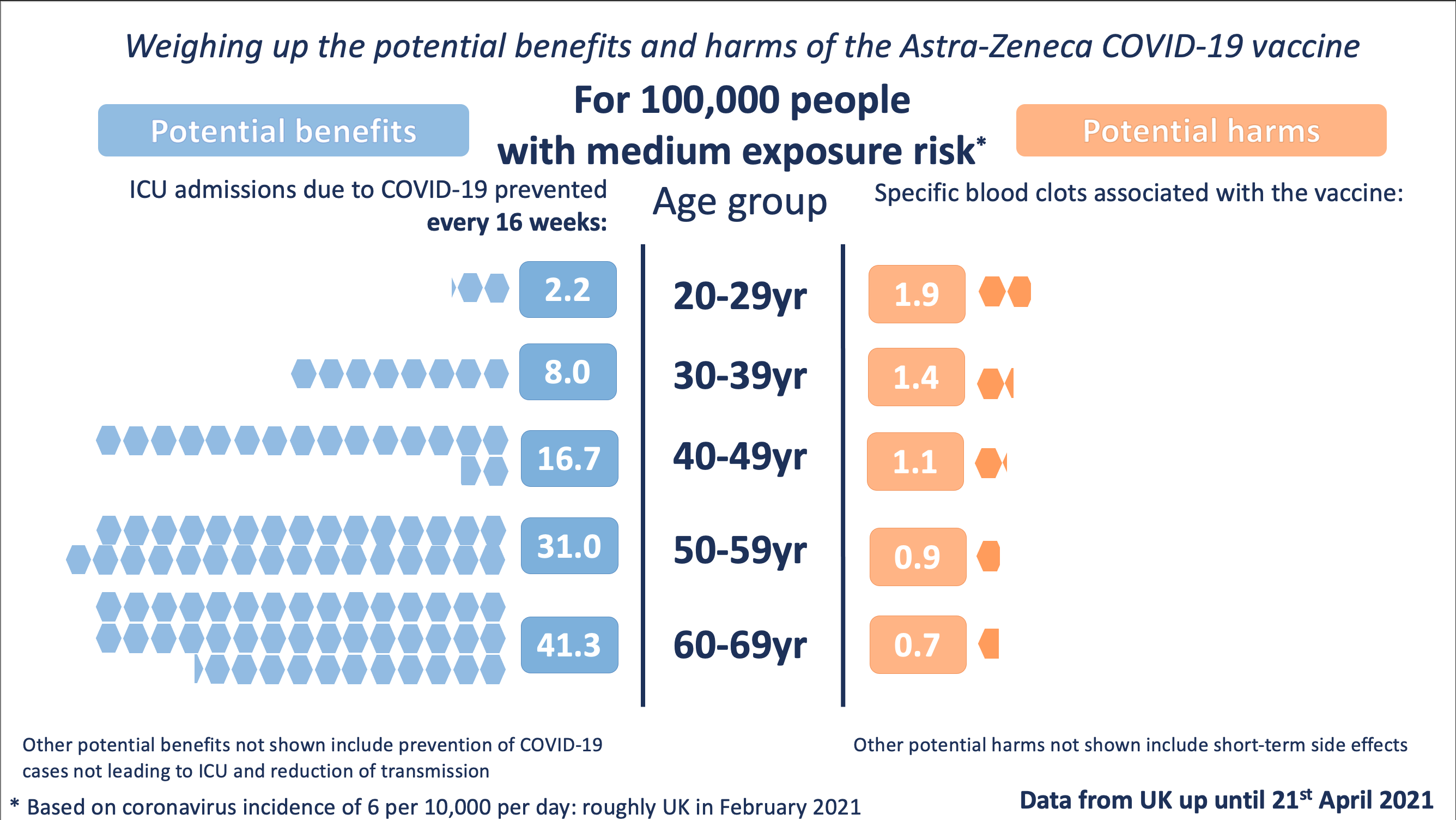
| From 2016 I have been Executive Director of the Winton Centre for Risk and Evidence Communication at the University of Cambridge. Evidence Communication is a way of trying to present trustworthy, useful information that other people can use to inform their decisions. Our motto is 'to inform, not persuade'. It's an interesting and different form of communication from many others. It is useful, for example, in producing information for patients facing healthcare choices (you can see some of the decision support tools we made for the NHS here. It is also useful in presenting evidence for policy-makers. For example, I helped produce graphics illustrating the potential benefits and harms of the Astra-Zeneca Covid-19 vaccine for different people and under different circumstances to communicate the first evidence from the MHRA to the JCVI. The Winton Centre brought together academic researchers, software engineers and professional communicators and we worked closely with graphics professionals. You can see my academic publications here. |
 |
 |
| In 2019 I set up my own not-for-profit company and went into partnership with JISC to produce Octopus, a new scholaraly publishing platform designed to help reset the incentive structure within academia (particularly STEM) towards intrinsic research quality. I won awards for its development from the Royal Society and Mozilla, and it is now funded by Research England and is in partnership with UKRN. |
 |
|
| Before 2016 I was a documentary film-maker, mainly for the BBC. I was Series Producer for BBC2's award-winning health/medicine magazine series Trust Me, I'm a Doctor, with presenters Michael Mosley, Gabriel Weston, Chris van Tulleken and Saleyha Ahsan. I also produced/directed many of the items, often self-shooting them, such as on ultrasound surgery or the world's first womb transplant. I also produced/directed a number of other programmes such as the first programme in the series Pain, Pus and Poison: The Search for Modern Medicines for BBC4 which went out in October 2013 (featuring the popular moment where Dr Michael Mosely tries the truth drug sodium thiopental). The programme won the 2014 'Learning on Screen' award. I also made Climate Change by Numbers, which won the AAAS Kavli Gold Award for Science Journalism.
|
|
|
|
| In the summer of 2012 I directed 4 of the BBC's 3D Olympic idents (Pole Vault, Long Jump, Taekwondo, Hurdling). This was a great project, in which we aimed to make the 3D as dramatic as possible, and really show off what it could do. The idents won an International 3D Society Award in 2013. The idents were filmed on 2 Phantom Flex cameras. The DoP was Ed Wild, and the stereographer Chris Parks. |
|
|
| Before then I worked on series including Bang Goes the Theory, for BBC1, Shock and Awe: The Story of Electricity for BBC4 and was Researcher or Assistant Producer on a range of BBC series such as The Making of Me; Walking with Beasts, the Walking with Dinosaurs Specials for BBC1; and Life in the Undergrowth with David Attenborough for BBC1. I also had my own format, Test Your Pet commissioned for BBC1. During this time I also produced websites to accompany such series (my Life of Mammals website won a Prix Europa and my Walking with Beasts website an RTS award, and The Big Al game, has had a life of its own! I won a BAFTA for my design for an interactive TV version of Walking with Beasts (featuring a 'scientific' alternative narration plus two extra video streams giving information alongside the main broadcast). |
|
Recent work:
| 1995-1998 | Linacre College, Oxford |
| D.Phil in animal behaviour with Dr. T. Guilford |
| Avaliable online here |
|
| 1992-1995 | New College, Oxford |
| MA degree in Zoology, with distinction in Supplementary Biological Anthropology. |
| Undergraduate project: "The morphology and behaviour of gorgonopsids and a new use for computers in palaeontology" available online here |
|
| 1985-1991 | Cheltenham Ladies' College |
|
| A-Levels: | Biology, Chemistry, Maths, Music
additional A-level courses taken in Further Maths, Art and Psychology (didn't feel the need to take more exams!) |
|
Education:
Other interests:
I just about find time outside of work to maintain my interest in music - singing locally with The Crown Singers and on an ad hoc basis with other groups. I also try to find time to shoot longbow regularly with The Companions of the Longbow.