
Figure 2-1 The Silver Washed Fritillary (Argynnis paphia): a) Male, b) Paphia female, c) Valesina female
This chapter investigates the basis of some interesting preferences found in a polymorphic butterfly species which appears to show a colour preference when choosing a mate and also the same colour preference when feeding from flowers. This species occurs in Britain with two population structures - one monomorphic and one polymorphic - thus presenting the opportunity to test the preferences of wild males in the two populations to see whether or not the males might be learning from their experience with the morph to which they are apparently not innately attracted. Also the presence of the same colour preference for mates and flowers is intriguing, suggesting that the males may possibly have a pre-existing bias for a colour which could have been exploited by the favoured female morph.
The Silver Washed Fritillary, Argynnis paphia, is a woodland butterfly which occurs in Britain, Europe, and across the Middle East to China (see Figure 2-1). The males and male-like females (morph paphia) are orange with black chequered markings. The sexes can be distinguished easily by the presence of black stripes on the males' forewings, which carry scent scales. The undersides of the wings are silvery green, making the butterflies well camouflaged when settled with their wings closed. There is a second female morph, valesina, which lacks the orange coloration. Instead, the upper surfaces of the wings are pale yellow or cream on the front of the forewings, changing to a dark green across the surface of the wings towards the abdomen. The frequency of valesina compared to the male-like morph, paphia, varies across the butterfly's range. In Britain valesina only occurs in central southern England, and then only at low frequency (about 5%). This frequency increases towards the Mediterranean and Far East, until the frequency of valesina apparently reaches 100% in Southern China (Ford, 1979 p168). Genetically, the alleles are autosomal: valesina is dominant, but sperm which carry the valesina gene appear to be at some disadvantage (Goldschmidt & Fischer, 1922).
Brunton et al. (1990) described valesina females in Corsica as approaching males, behaviour which was never seen from paphia females. This may be in order to allow the males to detect their sex pheromones if they are not visually attractive to males. It may also be normal for virgin females to show this behaviour, and that paphia females are usually mated quickly and so rarely demonstrate the approach behaviour, usually having been mated already. Brunton et al. also described how valesina females apparently had a higher heat tolerance, remaining at feeding sites during the midday heat whilst paphia females departed to the shade. This increased heat tolerance, and increased feeding times, may give the valesina females an advantage in hot countries. This increased advantage with increased temperature may explain, at least in part, the distribution of valesina, where the proportion of valesina increases approximately with increasing temperature, although the causal link is so far unexplained.
Magnus (1958) carried out an important and extensive series of experiments on the preferences of Argynnis paphia. Since these were originally published in German and have not been discussed widely in the English-language literature, I will report his findings here.
Using inexperienced butterflies he recorded their visits to paper flowers (repeating experiments carried out by Kühn and Ilse, 1925). He described carrying out these experiments at relatively low temperatures (between 15 and 20oC), and this meant that there was very little sexual activity (as measured by the frequency of the zig-zag flight characteristic of male mate-searching behaviour and approaches to other butterflies). Therefore he concluded that his results can safely be considered as flower colour preferences.
His results showed that both males and females show a preference for very bright orange/yellow (about PA3 in the Ostwald colour series), although the male peak is slightly higher and smoother, and pale blue (GA15 in the Ostwald series), where the female peak is higher. There is a lesser peak at pale green, which Magnus explained as being due to the butterflies searching for leaves on which to land in the cold.
Magnus carried out experiments on mate preference with wild butterflies. He found that there were no shape preferences and that males chose larger models over smaller models. He then investigated the attraction of moving models using an apparatus with a rotating arm in the field. He found a peak preference for orange (PA3-5), although there is also a lesser peak in the blue at PA13. Again he found that shape was not an important factor for the males and that attractiveness increased with size of the model. He then tested the effect of the model taking a flight path which fluctuated upwards and downwards (which he records as being characteristic of butterflies in 'feeding mode'), and found that models taking a straight path were preferred. Finally he tested the preferences of males for different frequencies of fluttering in the models. He found that higher frequencies of flutter were always preferred, up to the limit of the mechanism (40 cycles per second). He then continued the experiment with a new mechanism that spun a model around its axis of symmetry instead of trying to simulate actual fluttering. With this he recorded that males were increasingly attracted up to 50 revolutions per second. Above this they were not attracted, and he assumed this to be due to their flicker fusion frequency. Interestingly he noted that Hesperidae continued to be attracted at very high rates of revolution (sadly killing large numbers of them as they collided with the apparatus!)
Magnus then investigated the role of patterning in attracting males. He compared the normal female patterning to a model with no black at all, and one with denser, coarser, black markings and found that there was no significant difference between the attractiveness of the normal pattern and the model with no black at all, but that the model with more black was less favoured. He suggested that the males do not resolve the pattern at a distance, and that they rely on an 'overall effect' of colour, which was disrupted by the coarser black markings. He also tested the attractiveness of valesina against paphia females, and found that valesina was preferred only 10% of the time. However, he pointed out that the females release a pheromone which will trigger mating behaviour in the male in the absence of any colour stimulus, and so he concluded that the possession of this pheromone is what causes valesina females to be mated in the wild.
Interestingly, he then bred his own Argynnis paphia and tested the effects of genetics on the male preference for paphia females, and found that males homozygous and heterozygous for the valesina gene were indistinguishable from those homozygous for the paphia gene. This indicates that the gene, or gene complex, controlling male preference for either paphia or valesina females is not linked to the gene which causes the coloration itself. Therefore if a difference in the percentage of males with a preference for valesina is found between populations which varies in proportion to the percentage of valesina females in those populations, it cannot be ascribed to linkage disequalibrium between a gene complex controlling the coloration and one causing a preference for the coloration.
The results of the flower choice experiments, which showed that both inexperienced males and females had a preference for orange flowers (slightly greater in males than females) together with the results of the mate choice experiments, in which experienced males had a preference for orange mates (of any shape, preference increasing with size) suggests that both sexes have an innate preference for orange, and that this is being expressed in both flower choice and mate choice. However, since the males used in the mate choice experiment were wild, and therefore may well have experienced orange, paphia, females, it is also possible that they had learnt to recognise the common paphia females as mates. The 10% of males which preferred valesina in the choice experiments may correspond to those which had previously encountered valesina females (the proportion of valesina in the experimental population is not reported, although they were taken from Southern Germany where the proportion is said to be low (Brunton et al., 1990)). One way of testing the hypothesis that the preference is learnt is to repeat the mate choice experiments in two populations with varying proportions of valesina and paphia, such as in two areas of England, where it might be expected that the preference for valesina would vary with the proportion of valesina in the population.
The overall aim of the experiments is to determine whether male Argynnis paphia are innately attracted to orange, paphia, females - as they appear to be to orange flowers - or whether they learn a preference between the two morphs from their previous experience of females. Once this has been determined, the aim is to understand the basis for the mechanism and its implications for the evolution and maintenance of the valesina morph.
All experiments were carried out in Crab Wood, near Winchester, between the 10th July and 9th August, 1997. The butterflies were observed for about 30 hours during this period.
The butterflies spent the nights and cloudy periods resting up in trees, and when the sun emerged, would drop down like falling leaves into the sunny clearings and rides. Both sexes fly early in the morning (before 9am BST) if sunny. Females found bramble or thistle flowers (they were also seen on cow parsley), and fed whilst males patrolled the area, flying at the height of the bushy vegetation (mostly brambles and long grass). The males flew at other butterflies on the flowers, but appeared to be repelled by a sharp flash of the wings, at which they moved on. If a female didn't react in this way, the male would get underneath her and eventually push her off the flower. She then flew in a straight line with fluttering wings, with the male circling around her from front to back. If she then accepted him, the two started a 'wheeling' flight, both continually circling each other. Both sexes flew throughout the day whilst the weather remained very sunny, although the number of females seemed to decrease between about 11am and 3pm BST. Pairs were seen 'wheeling' throughout the day, until about 3:30pm BST, by which time there were very few Argynnis paphia around. Valesinas were seen at all times of the day, more often in the open, sunny coppiced areas. They were never seen to approach males as reported by Brunton et al. (1990), and one was seen to reject a male's advances. They were often approached by gatekeeper butterflies (Pyronia tithonus), which are about the same colour, but smaller. The proportion of valesinas was estimated to be about 7% of females through observation of females throughout the experimental period. About 150 females in total were seen during the 30 hours of observations, of which approximately 10 were valesinas. These butterflies were all thought to be different individuals, and the valesina females almost certainly were as seven of them were caught for use in the experiments and the variability in their coloration also made them individually recognisable.
Attempts were made to breed the species in captivity. The females laid eggs and small larvae emerged. In the wild these overwinter in the soil, but in captivity none re-emerged the following spring. In addition the adults proved difficult to keep alive, so all experiments were planned as fieldwork. The butterfly also appeared to be non-territorial, so repeated experiments on the same individual in the wild are not practical.
If the males of Argynnis paphia are initially attracted simply by colour, they might be expected to approach pinned-out males at least as often as paphia females. The males actually appear a brighter orange than the females, so they may approach the males more often than the females. If they approach the females more often, it is likely that they are detecting the female sex pheromone over a large enough distance to affect the results (since Magnus showed that the males showed no preferences between wing patternings). If they can detect the pheromones over such a distance, this would have to be taken into account when designing choice experiments. It is also possible that the males may approach other males in an aggressive manner (especially if a female is apparently present), so the behaviour of the males to both the pinned-out males and the paphia females was observed closely.
The aim of this experiment is to determine whether or not experienced, wild, males show a difference in their behaviour to pinned-out dead paphia females and the apparently brighter males, and also to establish the distance over which the female pheromones might be acting.
In order to test whether or not males discriminate between pinned-out males and pinned-out paphia females in their approach, a simultaneous presentation method was used, forcing the males to make a direct choice between a male and a female.
One dead male specimen and one dead paphia female specimen were presented in each experiment. The specimens were pinned through the thorax, with wings open, onto two corks held 50cm apart with wire, elevated to the height of the surrounding vegetation to maximise the chances of them being seen by free-flying males. These were placed in feeding areas where males were seen to be flying at the time of the experiment, and the specimens were positioned so as to be obvious from all angles of approach to avoid bias in the results due to the approach direction of the males. The apparatus was observed from a distance of a few metres away so as not to disturb the butterflies, and if a wild male was seen to make a deviation in his flight-path towards one of the pinned-out specimens it was recorded as a 'choice'. When the males landed on pinned-out specimens, attempts were sometimes made to mate, with both females and males. These were interrupted, however, by the removal of the male from the pinned-out specimen, as the fluttering of a male tended to attract other individuals in the area to investigate.
Once a male had been seen to make a choice between the two specimens, he was caught and kept in a mesh carrying-net for the rest of the day. If a male was not caught, the apparatus was moved 30-50m away for the next experiment to ensure that the data points were from different males and could thus be treated as independent as males might spend some time in each feeding-patch and thus their choices be recorded more than once. The time of each encounter, the first specimen the male approached, from which direction, and whether the male landed or attempted to copulate were all recorded.
Four male specimens and four paphia female specimens (collected from the area) were used in the experiment, and each possible combination of specimens was used twice - once with the male on the left and once with the female on the left All experiments were carried out in sunny weather between the hours of 9am and 4pm BST on 10th June, 22nd June, 23rd June, and 28th June 1997.
In most cases when a data point was recorded, the approaching male simply diverted his flight to fly around a pinned-out specimen briefly. In some cases, however, the male landed and showed considerable interest in the pinned-out specimen, which was noted.
| Male | Paphia female | |
|---|---|---|
| number of approaches | 24 | 8 |
| number of landings | 4 | 6 |
| percentage of approaches resulting in landing | 16.7 | 75 |
These results indicate that significantly more males approach pinned-out males than approach pinned-out paphia females in a simultaneous choice test (p<0.0035; binomial test; n=32).
A contingency table constructed for the result of the approaches (see Table 2-2) reveals that the 'expected landings' for the paphia female are too small to allow an accurate assessment of the difference between observed and expected for this cell, making a chi-squared test liable to overpredict the difference. However, when a chi-squared test is carried out, it is revealed that the chi-squared value (4.602) is large enough to give a p-value of less than 0.05 even when the value from this cell is entirely left out of the calculation (see Table 2-2). Therefore it can be concluded safely that the wild males landed significantly more often on the paphia females than the males once they had made an approach.
| approach to: | Male | Paphia female | |
|---|---|---|---|
| no landing | observed | 20 | 2 |
| expected | 16.5 | 5.5 | |
| landing | observed | 4 | 6 |
| expected | 7.5 | 2.5 | |
c2 = 0.742 + 2.227 + 1.633 + 4.9 = 9.503 ignoring value from cell with expected value less than 5, c2 = 0.742 + 2.227 + 1.633 = 4.602
Table 2-2 Contingency table constructed for observed and expected approaches and landing to dead males and paphia females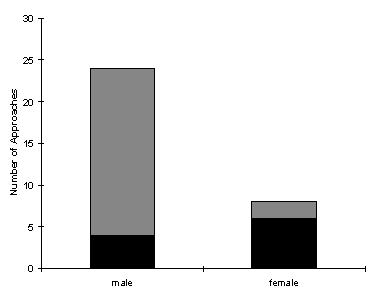
Many of the males approached from between the two pinned specimens, or it was not possible to determine from exactly which direction they approached. Therefore, only a small amount of data on approach direction was collected. However, as the figures in Table 2-3 suggest, the direction from which males approached appeared not to affect their choice.
| Approached from right | Approached from left | Approached perpendicular to experiment | |
|---|---|---|---|
| Chose right hand specimen | 3 | 5 | 4 |
| Chose left hand specimen | 1 | 2 | 2 |
In general, then, it can be concluded that males are more frequently attracted to pinned-out male specimens than pinned-out paphia females, but that their reaction once they had approached was different. When females were approached, the males landed (presumably to attempt copulation) significantly more often than when they approached males. Males were attracted to other patches of bright orange (such as car indicator lights), but when attempts were made to pin out painted paper models, not one male showed any response. This supports the results of Magnus (1958), that males are attracted to very bright orange, but that this attraction is lost as the orange colour pales (the painted models were made with water colour paint on white paper, so the orange colour had a relatively low saturation). However, at close quarters the males appear to be able to differentiate between the sexes, as they land on the females significantly more frequently. This again accords with Magnus' results, as at very close range the males can detect residual pheromones on the females, which causes the males to attempt copulation. Magnus found the colour PA4 in the Ostwald colour series to be the most attractive to the males, and showed that saturation of the colour was very important (less saturated oranges quickly lost their attractiveness). PA4 was the most saturated orange colour. Although it has not been possible to determine the spectrum of the PA4 paper, spectra could be obtained for the ground orange colour in the males and females of Argynnis paphia to confirm the apparent greater brightness (saturation) of the male colouring. This is done in Experiment 2-3.
Since Experiment 2-1 demonstrated that (outside a very close distance) males follow visual rather than pheromonal attraction clues, the experimental procedure was considered suitable for testing the relative attractiveness of paphia and valesina females to males of differing experiences.
Magnus' results (1958) together with Experiment 2-1 suggested that males might be attracted to potential mates simply through their orange coloration. However, Experiment 2-1 also indicated that at close range males can recognise mates; this is probably through the detection of pheromones, which would allow males to recognise valesina females as mates despite their coloration lacking the attractiveness of orange. Thus valesina females are usually mated in the wild, and it is possible that once a male has mated with a valesina female he might learn to recognise the valesina coloration despite the lack of an innate preference. If males are learning to recognise mates through their previous experience, then experienced males might be expected to show a preference for valesina related to the frequency of valesina in the population. The possibility that the same effect might be due to a genetic factor has already been ruled out by Magnus.
In order to test this it should be possible to carry out a choice experiment in both an area where valesina is present, and an area where valesina is absent. If a preference for valesina in some males is found in the area where they are present which is not found in the area where valesina is absent, it seems likely that learning might be occurring. The experiment could be carried out in England because valesina is present (in a low proportion) in central southern regions, and completely absent from other populations elsewhere in the country.
The aim of this experiment is to determine whether or not males show a preference for valesina females related to the frequency of valesina in the area.
The same simultaneous choice procedure was used as in Experiment 2-1, with one dead paphia female specimen and one dead valesina female specimen being presented pinned out on corks held 50cm apart. The specimens were again raised to the level of the surrounding vegetation and placed in feeding areas where males were most likely to notice them. The choices of approaching males was recorded in the same way, recording the time of approach and the male's behaviour (whether he landed or attempted to copulate). As before, males were caught after each approach, and kept in carrying-nets to prevent the choice of a male being recorded more than once. If a male was not caught, the apparatus was moved 30-50m away from the previous site.
Five dead specimens each of paphia and valesina females, caught in the area, were used in rotation. The females displayed varying wing conditions, but there was no difference in general between the conditions of the paphia females and that of the valesina females. Experiments 2-1 and 2-2 were run concurrently, with both sets of apparatus being set up and observed in the same area on some days. The trials were all carried out in Crab Wood, near Winchester, on 22nd July, 23rd July, 28th July, and 9th August.
| Number of approaches to paphia female | Number of approaches to valesina female |
|---|---|
| 25 | 8 |
These results indicate that males significantly prefer paphia females to valesina females (p=0.0023; binomial test, n=33). When the results were examined closely, however, those relating to one individual valesina appeared anomalous. Seven out of the eight approaches to valesina females were to this one particular individual (number 3), and this female was preferred in 7 out of 10 of the trials in which she was used. Moreover, in all but one case, the males who chose this female landed on her, and this behaviour was similar to that seen when a female which had been freshly killed by being squeezed tightly was pinned out (which was done in the initial pilot tests). It seems likely, therefore, that this individual carried a particularly large amount of attractive pheromone (possibly she was a virgin) and that this affected the results of the experiment, whereas the other females had all been mated and carried lesser amounts of pheromone which wore off between their being killed and being used in the experiments. It might be argued that the five paphia females might also be carrying large amounts of pheromones, thus explaining their attractiveness to males compared with the other valesina specimens. However, firstly they were all found to be less attractive than valesina 3 in pairwise tests. Secondly, if one 'particularly attractive' valesina was found out of five, and the probability of finding an equally attractive paphia was the same, then the probability of all five paphia specimens used being 'particularly attractive' is 0.00032. Thus this can be seen to be very unlikely.
Table 2-5 shows the data if all the results of experiments involving valesina specimen 3 are removed:
| Number of approaches to paphia female | Number of approaches to valesina female |
|---|---|
| 22 | 1 |
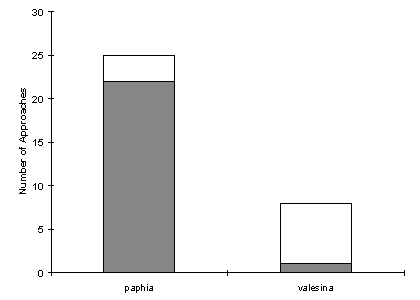
This data gives an even stronger bias towards paphia (p=3x10-6, binomial test, n=23), showing that the males approach paphia females significantly more frequently than valesina females.
In addition, the data for the experiment involving two female morphs was obtained much more slowly than that for the experiment involving a male and a female. Since the two experiments were run concurrently in the same area for some of the time, the rate of acquisition per hour can be analysed:
| day | hour (BST) | male vs. female | valesina vs. paphia |
|---|---|---|---|
| 1 | 12-1 | 1 | 2 |
| 10-11 | 3 | 0 | |
| 11-12 | 3 | 1 | |
| 2 | 12-1 | 6 | 3 |
| 1-2 | 3 | 0 | |
| 2-3 | 1 | 0 | |
| 3-4 | 5 | 1 | |
| 9-10 | 1 | 0 | |
| 3 | 10-11 | 4 | 2 |
| 11-11:30 | 1 | 1 | |
| TOTAL | 28 | 10 | |
The difference between the total amount of data recorded for each experiment during these time periods indicates that the male vs. paphia experiment attracted significantly more males than did the valesina vs. paphia experiment (p=0.0025, binomial test, n=38).
The results of Experiments 2-1 and 2-2 accord with those of Magnus (1958), showing that males seem to be attracted to the bright orange of pinned-out males and to other orange objects, although they do recognise females at close quarters as they land more often on the females than on the males, probably due to residual pheromones rather than visual recognition. This attraction for orange would explain why paphia females seem significantly more attractive to the males than the valesina females, which they rarely appear to recognise as mates at a distance. However, the data from valesina 3 might suggest that the valesina females carry the normal sex pheromone which makes them attractive to the males when the males detect the pheromone (as was postulated by Magnus). The lack of visual recognition may put the valesina at a disadvantage when they are at a distance from the males, but where males are finding females at relatively close quarters, such as at a feeding site (or if valesina females actively approach males), the pheromones are more important (as Magnus describes mating attempts occurring in the absence of any visual signals when the pheromone is present). Cook et al. (1994) suggested that female morphs which appeared not to be attractive to males at a distance may actually be at an advantage in avoiding 'sexual harassment' which could result in wing damage and lost feeding time etc. Female Argynnis paphia however were never seen to be 'harassed' unduly in the wild, and seemed to be able to feed alongside males without trouble. It does not appear, therefore, that valesina females would be at an advantage by avoiding male attentions.
Experiment 2-2 (after the removal of the anomalous data from valesina specimen 3) showed only one male out of 23 approaching a valesina female in a choice between valesina and paphia. The hypothesis that males may learn to recognise (and even prefer) valesina females as mates after an initial mating with a valesina female remains tenable, but could not be demonstrated by fieldwork in this country. Experiment 2-2, carried out in an area where valesina occurs, showed such a small preference for valesina that if the experiment were to be repeated in an area where valesina did not occur and no males approached the valesina (as would be the extreme of the hypothesis - that males only recognise valesina females as a mate after first mating with one) there would be no significant difference between the data sets. Therefore to test the hypothesis, the experiment would have to be repeated in an area where valesina occurs at a much higher frequency to determine whether or not the preference for valesina illustrated by the males increases with it. Thus it has not been possible to test for the affect of learning on the males' choice of mates.
It is interesting that that the males appear to show identical colour preferences when searching for food and mates (the difference in their flight patterns recorded by Magnus indicating that they were showing two different behaviours). Such a preference could be due to a peak in their visual sensitivity, or it could be an innate preference which has either evolved to allow the males to recognise their mates (and/or food) or which is just the result of random drift resulting in a 'perceptual bias'. In order to clarify this area in Experiment 2-3 the colours of the male and females are compared, and in Experiment 2-4, the visual sensitivities of the butterflies are explored.
Male Argynnis paphia show a preference for males over paphia females, and paphia females over valesina females in choice tests (Experiments 2-1 and 2-2). This preference ranking appears to be due to the differing amount of orange coloration found in the males and two morphs (Magnus 1958). The paphia females of Argynnis paphia appear to have a slightly less saturated orange background colour than the males (and the valesina females are greenish rather than orange), although it is not possible to quantify this without measuring the reflectance spectra of both male and female wings. Magnus' experiments indicated that the increased black patterning on the females' wings did not make a difference to the preferences of the males, so Experiment 2-3 is designed to quantify the orange ground colour in between the black markings of either sex.
The aim of this experiment is to quantify the apparent difference in saturation between the orange colour of the male and paphia females of Argynnis paphia.
A Zeiss MC5-501 UV-Vis spectroradiometer was used to measure the reflectance spectrum of the butterfly wings. The wing was illuminated with light from a xenon bulb at 45o to the normal, with measurement of the spectrum taken normal to the surface (with no integration of the scattered angular reflection). The measurements were taken of a circular area approximately 1mm in diameter. Measurements were taken of the background colour of the forewings of two specimens of either sex, and of the hindwings of two specimens of either sex. For comparison, the background colour of the forewing of one specimen of the valesina morph was also measured. The area measured was the same in each individual, and was chosen to include as little as possible of the black markings. The instrument was calibrated on an artificial true-white Spectralon( disc, against which all the reflectivities were measured.
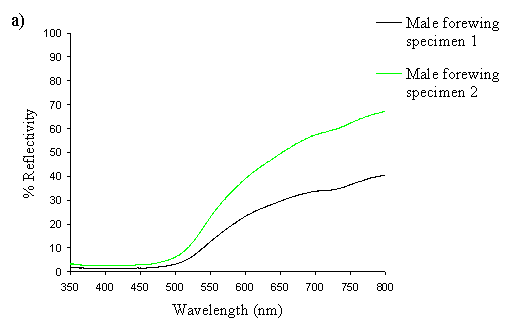
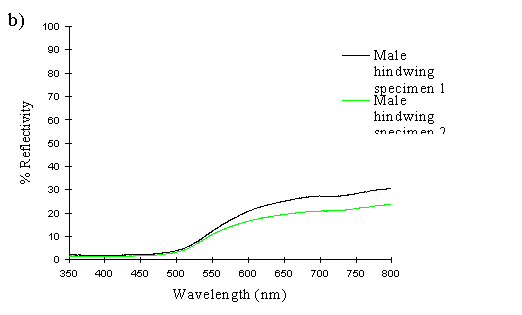
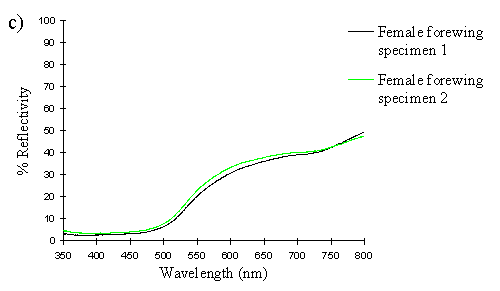
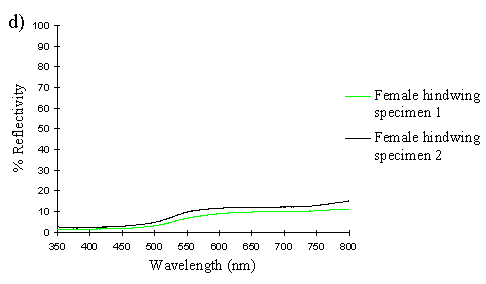
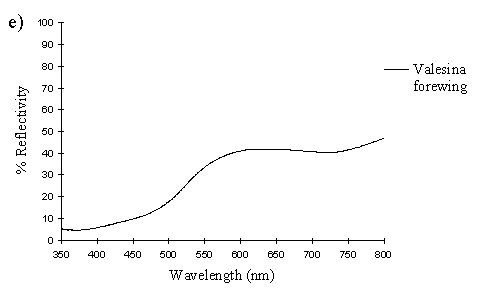
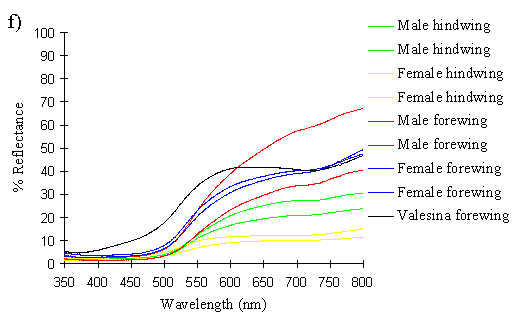
The spectra clearly show several things. Firstly, the male and paphia female wings all show the same shape spectrum, indicating that they are essentially the same colour (hue). They all show a smooth curved decline in reflectance from 750nm to around 500nm, resulting in an orange coloration (with no UV reflectance). The spectrum of the valesina female forewing is a slightly different shape, showing a relatively constant level of reflectance from 750nm to 550nm, followed by a sharper decline in reflectance to 500nm. The reflectance between 500nm and 350nm in the valesina female is higher than in the males and paphia females.
Secondly, the forewings of males and paphia females show a higher reflectance than their respective hindwings, indicating a brighter colour (higher saturation). Thirdly, both the forewings and hindwings of the males are more reflectant than the forewings and hindwings of paphia females respectively, showing that the males are indeed a brighter (more saturated) orange than the females. Although one of the male forewings appears to be less saturated than both the female forewings, the two are, on average, more saturated than the two female forewings. It is possible that this uncharacteristic difference in the reflectivities of two individuals is due to a small amount of black patterning being included in the area analysed, decreasing the overall intensity.
These results therefore confirm that the males are a brighter orange than the paphia females, but shows that they are fundamentally the same colour (hue). Magnus (1958) showed that male Argynnis paphia were most attracted to the colour PA4 in the Ostwald colour system - the most saturated orange colour - and were less attracted to the less saturated oranges of the same hue (LA4, GA4). This is consistent, therefore with the results of this experiment and those of Experiment 2-1, where males were shown to be more visually attracted to other males than to paphia females.
Experiments 2-1 and 2-2, and the work of Magnus (1958) indicate that the males have a preference for bright orange coloration, and this appears to be manifested in both their mate choice and choice of flowers for feeding. This preference could be due to an increased visual sensitivity for these wavelengths of light, or a perceptual bias for this colour (which itself could either have evolved in order to allow males to detect females easily, or could be selectively neutral). It is possible to assess objectively the spectral sensitivities of animals, and thus determine whether a preference is due to the actual reception of the signal or to a bias in the processing of the signal received. Electroretinograms (ERGs) can be used to measure the neural response of an individual to visual stimuli, and have been used to assess the colour sensitivities of many Lepidoptera by recording the magnitude of the electrical response of the retina to flashes of light of differing wavelengths:
| Species | 1o peak (nm) | 2o peak (nm) | 3o peak (nm) | 4o peak (nm) | Reference |
|---|---|---|---|---|---|
| Atrophaneura alcinous | 460 | 420 | 520 | 600 | Eguchi et al (1981) |
| Papilio machaon | <380 | 460 | 520 | 580 | Eguchi et al (1981) |
| Papilio xuthus | <380 | 480 | 520 | 580 | Eguchi et al (1981) |
| 400 | 460 | 360 | 520 | Arikawa et al. (1987) | |
| Papilio protenor | 420 | 460 | 520 | Eguchi et al (1981) | |
| Papilio bianor | 460 | 520 | 420 | 580 | Eguchi et al (1981) |
| Papilio maaki | <380 | 460 | 520 | 580 | Eguchi et al (1981) |
| Papilio troilus | 483 | 578 | (UV not studied) | Swihart (1970) | |
| Graphium sarpedon | 560 | 600 | <380 | 460 | Eguchi et al (1981) |
| Colias erate | 400 | 520 | 560 | Eguchi et al (1981) | |
| Gonepteryx aspasia | <380 | 460 | 560 | 620 | Eguchi et al (1981) |
| Pieris melete | 400 | 480 | 600 | 540 | Eguchi et al (1981) |
| Lycaena phlaea | 400 | 600 | 540 | Eguchi et al (1981) | |
| Lycaena rubidus & Lycaena heteronea | 360 | 437 | 500 | 568 | Bernard & Remington (1991) - distribution varies throughout eye. |
| Pseudozizeeria maha | 400 | 580 | 520 | Eguchi et al (1981) | |
| Celastrina argiolus | <380 | 440 | 560 | Eguchi et al (1981) | |
| Parantica sita | 520 | <380 | Eguchi et al (1981) | ||
| Argyronome ruslana | 560 | <380 | 620 | 440 | Eguchi et al (1981) |
| Polygonia c-album | 560 | 600 | <380 | 520 | Eguchi et al (1981) |
| Inachis io | 560 | <380 | 520 | 460 | Eguchi et al (1981) |
| Aglais urticae | 360 | 460 | 530 | Scherer & Kolb (1987) | |
| Parage aegeria | 360 | 460 | 530 | Scherer & Kolb (1987) | |
| Fabriciana adippe | 580 | 520 | 460 | <380 | Eguchi et al (1981) |
| Nymphalis xanthomelos | 560 | <380 | 500 | 460 | Eguchi et al (1981) |
| Minois dryas | 520 | <380 | 460 | Eguchi et al (1981) | |
| Neope goschkevitschii | <380 | 520 | 460 | Eguchi et al (1981) | |
| Ochlodes venata | 520 | 460 | 380 | Eguchi et al (1981) | |
| Parnara guttata | 520 | 460 | <380 | Eguchi et al (1981) | |
| Cephonodes hylas hylas | 520 | 460 | <380 | Eguchi et al (1981) | |
| Marumba sperchius sperchius | 540 | 460 | 600 | Eguchi et al (1981) | |
| Ampelophaga rubiginosa rubiginosa | 540 | 460 | 580 | Eguchi et al (1981) | |
| Callambulyx tatarinovii japonica | 540 | <380 | 460 | 580 | Eguchi et al (1981) |
| Macroglossum pyrrhostictum | 500 | 460 | Eguchi et al (1981) | ||
| Anadevidia peponis | 460 | 500 | 420 | Eguchi et al (1981) | |
| Mamestra brassicae | 460 | 580 | 560 | <380 | Eguchi et al (1981) |
| Samia cynthia ricini | 530 | 480 | 580 | <380 | Eguchi et al (1981) |
| Actias artemis aliena | 540 | <380 | 460 | 580 | Eguchi et al (1981) |
| Phassus excrescens | 440 | 400 | 520 | <380 | Eguchi et al (1981) |
| Arichanna gaschkevitschii gaschkevitschii | 540 | 500 | <380 | Eguchi et al (1981) | |
| Epicopeia hainesii hainesii | <380 | 420 | 500 | Eguchi et al (1981) |
These measurements were all taken with light of the same intensity (quantum flux), and the resulting graphs of electrical response of the retina against wavelength of light are known as spectral efficiency functions (Menzel, 1979 p507). However, as is pointed out by Menzel, the response-intensity function at different wavelengths can be very variable, and so an animal may seem to have a peak sensitivity to one wavelength at one intensity level may have a different peak sensitivity at another intensity level. Hence, a 'surface' of sensitivity to different wavelengths at different intensities is most meaningful, although the values at the lowest end of the intensity range are probably the most representative of the intensities reflected off surfaces (such as flowers and other butterfly wings) so the values in the table above probably give a reasonable view of the visual capabilities of the insects.
Most ERG experiments have used a 500W xenon lamp as a light source (the beam of which is then passed through a monochromator). Xenon lamps have a relatively even but fairly low intensity across the spectrum, and the quantum flux in most experiments was brought down to the level of the lowest intensity wavelength by using filters. This ensures an equal but low intensity from which to make the response measurements. Measurements of responses to higher intensities of light appear not to have been made, and a 'response surface' has not been constructed.
The aim of the electroretinogram is to determine whether the colour preferences of the butterflies can be explained by peaks in their visual sensitivities.
The wings of the butterfly were removed, and the butterfly was placed in a tightly fitting plastic tube, with only the head protruding. The legs and antennae were prevented from moving by the tightness of the tube. Two thin silver wire electrodes were placed just under the surface of one of the eyes, one near the dorsal surface of the eye and one nearer the ventral surface. The impulses from the eye were amplified using an Isleworth preamplifier (amplified x1000, no low or high pass filtering), and recorded on a Tektronix Type 502A dual-beam oscilloscope and a Lloyd Instruments PL3 chart recorder. The tube containing the butterfly was placed in an earthed metal Faraday cage to shield the electrodes from surrounding electrical appliances. The light source used was a 200W mercury lamp (with an infra-red filter in the form of a lens containing continuously flowing water to prevent overheating) and a Spectral Energy GM252 monochromator, focused by a silicon lens onto the eye, and this was the only light incident on the eye within the metal box (see Figure 2-5). The light beam was interrupted by card between the IR filter and the monochromator until a steady background signal from the eye was obtained, also dark-adapting the eyes. The card was then sharply removed and immediately replaced, producing a spike of electrical activity from the retina.
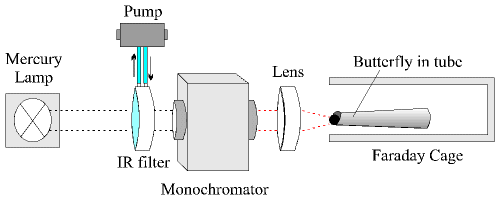
Single measurements were taken in this way at 25nm intervals through the spectrum from 250nm (where no response was found) to 850nm or vice versa. These 'runs' were repeated about 5 or 6 times on each individual butterfly, and the procedure was repeated for 6 individuals. Four females and two males were used (no more male specimens were available). However, no sex differences have been reported previously, and so a difference in the visual sensitivities of the sexes was not expected.
This procedure gave the responses of the retina to light of the highest intensity which could be produced by the mercury lamp (much higher than a corresponding xenon lamp). However, the intensity of a mercury lamp is by no means constant across the spectrum, so this was measured using a photodiode, RS stock number 305-462 (RS Electronics*) (see Appendix 2 for further details). A set of filters (numerical swatch set, Lee Filters**) was then calibrated using the photodiode at the peak wavelengths of the lamp (375nm, 450nm, 550nm), which also correspond roughly to the peak sensitivities of the three photopigments found in Hymenoptera (Chittka & Menzel, 1992; Backhaus & Menzel, 1992), Manduca sexta (Bennett et al., 1997), three of the four pigments found in Lycaena buttterflies (Bernard & Remington, 1991), and three of the five pigments found in Papilio xuthus (Arikawa et al., 1987). The response of four individual butterflies was thus recorded in the same way to differing intensities of these three wavelengths. Since these showed that there was little or no variation in the intensity functions of different individuals, and that the data formed smooth curves with little noise, the surface was constructed using multiple readings from only one individual. The advantage of this was that the butterfly was not moved throughout the series of readings, and so the same proportion of the beam was falling on the eye at all times from the lamp. If different individuals had been used and placed in the beam, then inevitably some would have received slightly more or slightly less of the beam's intensity, and therefore the readings from different individuals would all be slightly shifted up or down the intensity curve relative to each other (see Appendix 2 for full details).
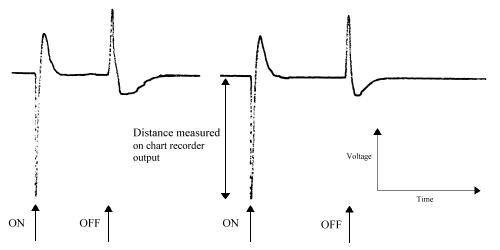
The wave-form of a typical lepidopteran ERG is described as a monophasic negative response (Eguchi et al., 1982). This is seen as a characteristic 'spike' above background noise in response to light stimulation (see Figure 2-6). The spikes of activity from the eye are quite difficult to analyse as occasionally an initial positive response is seen which is a response from the optical ganglion layers (Swihart & Gordon, 1971; Eguchi et al., 1982). Where this occurred, the positive response was ignored.
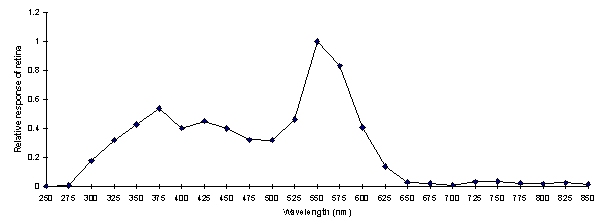
All the individuals showed a very similar spectrum of sensitivity to wavelength, but on varying scales of amplitude. The individual spectra were therefore normalised with respect to each other. This was done by selecting one spectrum (the one in which the component 'runs' were most similar), and calculating the ratio of each point in an individual spectrum to the respective point in the selected spectrum. The mean ratio between each spectrum and the selected spectrum could then be calculated. Each point in each individual spectrum was then multiplied by this mean ratio for the particular spectrum to scale all the spectra with respect to that selected. Then a mean spectrum for all individuals was calculated. To avoid biasing the result due to the initial selection of one spectrum to which to normalise all the others, this mean spectrum was then used as the 'selected spectrum' and the process iterated until a converged mean spectrum was achieved (see Figure 2-7).
This spectrum was then calibrated for lamp intensity. For this, measurements from another species, Papilio dardanus, had to be used due to the unavailability of further Argynnis paphia. Although this is not ideal it is justifiable, as the similarity between the two uncalibrated response curves (see Experiment 3.5 for efficiency spectrum of Papilio dardanus) suggests that the two species are similar in their responses to different wavelengths and intensities. See Appendix 2 for further details on the calibration of the spectrum.
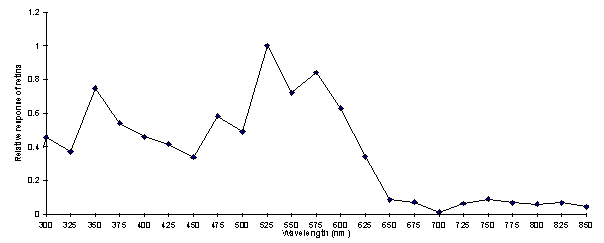
Although the spectral efficiency function may be slightly biased by the fact that the intensity spectra used were measured from Papilio dardanus, it is probably a fair estimate of the visual capabilities of the species. The three main visual pigments found in insects are present, at approximately 360nm (350nm peak), 460nm (475nm peak), and 520nm (525nm peak) (Chittka & Menzel, 1992; Backhaus & Menzel, 1992; Bennett et al., 1997; Bernard & Remington, 1991), with another, slightly broader peak at 575nm similar to that found in Papilio dardanus (Experiment 5-3), Papilio xuthus (Arikawa et al., 1987), Papilio aegeus (Matic, 1983), Papilio troilus (Swihart, 1970) and others in Table 2-7).
The presence of response to wavelengths above 700nm is intriguing. The response is very small compared with that found between 300 and 650nm, but all the same it was quite marked in many individuals, whilst extinction of the response was achieved at 250nm and below (see Figure 2-7 - the response below 300nm is not shown in Figure 2-8 as the sensitivity of the light meter was too low at these wavelengths to produce a meaningful calibration). Above 700nm, the structure of the eye should filter out most if not all of the incident light, and thus render detection by the animal nearly impossible. In addition, photopigments that respond to such wavelengths are very rare (although Douglas et al. 1998 have identified some chlorophyll derivatives such as pheophorbide a and pyropheophorbide a which have absorption spectra around 700nm and above, and have shown their use in the dragon fish Malacosteus niger). It seems unlikely that the butterflies would indeed be sensitive to this region of the spectrum, and this result could simply be due to a fault in the particular monochromator used which let light of wavelengths other than 750-850nm through, but no others were available to repeat the experiment. It should also be noted that the when the monochromator was set to wavelengths between 750-850nm no light of other wavelengths visible to the human eye were seen to be passing through.
Comparing this spectrum of sensitivity with the reflectance spectra of the wing colours of the species it can be seen that the preference for the orange of males and paphia females over the greener coloration of the valesina females cannot be ascribed to a visual bias for the colour orange. In fact, the valesina females might be expected to be more attractive as they have a higher reflectance around 525nm, the peak sensitivity for the species. Therefore the preferences shown by the males must be due to higher neural processing.
Male Argynnis paphia appear to be attracted to highly saturated orange coloration, both when choosing mates and flowers (Magnus, 1958). Their preference for the most highly saturated orange accords with them being more attracted to the brighter orange males than the less highly saturated orange paphia females (results of Experiment 2-1), and not visually attracted to the greener valesina females (results of Experiment 2-2). The apparent difference in saturation between the males and the paphia females has been confirmed by reflectance spectroscopy (Experiment 2-3).
This preference appears not to be due simply to a visual sensitivity bias in the retina of the butterfly (results of Experiment 2-4), and so seems to be due to the perception rather than the reception of the signal. This preference must either have been selected for in evolution or be due to random, unselected drift.
If the preference were initially due to random drift, it could have then been exploited by the paphia females in order to increase their visual attractiveness to males. The fact that they are not as bright (and visually attractive to males) as the males could be due to a trade-off between this and the need to be cryptic, in order to lay as many eggs as possible, although it is unclear why in this case the males should have evolved an orange coloration (and one which is slightly different from that of the females). The fact that Magnus (1958) showed a lesser reaction to the bright orange in the females indicates that the roles are unlikely to be reversed - with the males becoming brighter in order to attract females. The sex difference in perception also suggests that the preference may have been selected for in the males. If this were the case, the bias would have evolved as those individuals who reacted to orange stimuli particularly strongly achieved more successful matings with the orange females and left more offspring than those individuals who did not.
It is not easy to differentiate between the adaptive and random drift possibilities, nor to determine whether or not the females are showing sensory exploitation of the males. It would be very interesting to repeat the colour preference experiment with males from an area where valesina is the most common form of the female (such as in Southern China) to determine whether or not the males retain their preference for orange paphia females, or whether they have an innate preference for the valesina coloration. If they still retain the, now maladaptive, preference for orange mates then it would seem more likely that the preference is due to random drift. It would also be interesting to look at the phylogeny of the colour preference, to determine whether it is shown in other related fritillaries (especially those where the ground colour is not the same bright orange).
Since Magnus' results show that the males show some interest in blue models, it might be possible for a blue female morph to exploit this attraction. In this species it seems unlikely that this would occur as the females must remain cryptic. However, it is be possible that such a bias might be exploited in a polymorphic mimetic butterfly such as Papilio dardanus. Therefore the flower colour preferences of Papilio dardanus are investigated in Chapter 4, in order to compare these with the mate colour preferences found by Cook et al. (1994), and also further investigated in Chapter 5.
I would like to thank Hampshire County Council for allowing me to carry out the field work in Crab Wood. I would also like to thank Innes Cuthill for the use of the spectroradiometer, and Richard Compton for the use of the xenon lamp, monochromator and chart recorder. I am deeply indebted to Frank Marken for help with translations from German, and to Steve Simpson and John Alden for help with the ERG.
Arikawa, K., Inokuma, K., Eguchi, E. 1987. Pentachromatic visual system in a butterfly. Naturwissenschaften 74, 297-298.
Backhaus, W. & Menzel, R. 1992. Conclusions from the colour vision of insects. Behavioral and Brain Sciences 15 (1), 28-30.
Bennett, R.R., White, R.H., Meadows, J. 1997. Regional specialisation in the eye of the sphingid moth Manduca sexta: blue sensitivity of the ventral retina. Visual Neuroscience 14, 523-526.
Bernard, G.D. & Remington, C.L. 1991. Color vision in Lycaena butterflies: spectral tuning of receptor arrays in relation to behavioural ecology. Proc. Natl. Acad. Sci. USA 88, 2783-2787.
Brunton, C.F.A., Baxter, J.D., Quartson, J.A.S., & Panchen, A.L. 1990. The valesina morph of the butterfly Argynnis paphia (L.) in Corsica, 1988. Ent. Rec. 102 (1-2), 31-7.
Chittka, L. & Menzel, R. 1992. The evolutionary adaptation of flower colours and the insect pollinators' colour vision. J. Comp. Physiol. A 171, 171-181.
Cook, S.E., Vernon, J.G., Bateson, M. & Guilford, T. 1994. Mate choice in the polymorphic African swallowtail butterfly, Papilio dardanus: male-like females may avoid sexual harassment. Anim. Behav. 47, 389-397.
Douglas, R.H., Partridge, J.C., Dulai, K., Hunt, D., Mullineaux, C.W., Tauber, A.Y., and Hynninen, P.H. 1998. Dragon fish see using chlorophyll. Nature 393, 423-4.
Eguchi, E., Watanabe, K., Hariyama, T., Yamamoto, K. 1981. A comparison of electrophysiologically determined spectral responses in 35 species of Lepidoptera. J. Insect Physiol. 28 (8), 675-682.
Ford, E.B. 1979. Ecological Genetics. Chapman & Hall, 166-8.
Goldschmidt, R. & Fischer, E. 1922. Argynnis paphia-valesina, ein fall geschlechtskotrollierter Verebung bei Schmetterlingen. Genetica 4, 247-278.
Kühn, A. & Ilse, D. 1925. Die Anlockung von Tagfaltern durch Pigmentfarben. Biol. Ztrbl. 45, 144-149.
Magnus, D.B.E. 1958. Experimentelle Untersuchungen zur Bionomie und Ethologie des Kaisermantels. Z. Tierpsychol. 15, 397-426.
Mati(, T. 1983. Electrical inhibition in the retina of the butterfly Papilio. Four types of photoreceptors. J. Comp. Physiol. 152, 169-182.
Menzel, R. 1979. Spectral sensitivity and color vision in invertebrates in Comparative physiology and evolution of vision in invertebrates ed. Autrum, H. & Myrick, E. (Handbook of sensory physiology v. VII/6A), Springer-Verlag, Berlin, 503-580.
Menzel, R. 1985. Colour pathways and colour vision in the honeybee in Central and peripheral mechanisms of colour vision ed. David Ottoson & Semir Zeki (Wenner-Gren Center international symposium series; v.43), 211-233.
Scherer, C. & Kolb, G. 1987. The influence of color stimuli on visually controlled behavior in Aglais urticae (L) and Parage aegeria (L) (Lepidoptera). J. Comp. Physiol. A 161 (6), 891-898.
Swihart, S.L. 1970. The neural basis of colour vision in the butterfly Papilio troilus. J. Insect Physiol. 16, 1623-1636.
Swihart, S.L. & Gordon, W.C. 1971. Red photoreceptor in butterflies. Nature 231, 126-127.
|
Please cite this thesis as: Freeman, ALJ; 1998; D.Phil thesis, Oxford University. |
E-mail to Alexandra Freeman Back to Table of Contents Back to Home Page |