
Figure 4-1 A male Papilio dardanus feeding off a blue artificial flower in Experiment 4-1
There is a wide range of flowers available to nectar-feeding insects, and the range changes with time. Studies on bees (von Frisch, 1914) first showed that such insects can show innate colour preferences which can subsequently be changed by learning, and similar experiments on various butterfly species have shown that they also have innate preferences (see Table 4-1), and show a considerable degree of flower constancy (Goulson & Cory, 1993).
| Species | Colour preference | Reference |
|---|---|---|
| Heliconius charitonius | primary: orange/red secondary: blue/green | Swihart & Swihart (1970) Swihart (1971) |
| Battus philenor | primary: yellow secondary: blue/purple | Weiss (1997) |
| Aglais urticae | blue/green & yellow/red primary: 460nm secondary: 582nm | Ilse (1928) Scherer & Kolb (1987) |
| Pararge aegeria | primary: 450nm secondary: 630nm | Scherer & Kolb (1987) |
| Heliconius erato hydara | yellow/orange | Crane (1955) |
| 'tropical Papilionids and Pierids' | red (observation in field only) | Crane (1955) |
| Pieris brassicae | orange/red & blue/purple primary: 447nm secondary: 600nm | Ilse (1928) Scherer & Kolb (1987) |
| Pieris rapae | primary: purple/blue secondary: yellow Some individuals flower-constant, whilst others not. | Kandori & Ohsaki (1996) |
| Pieris napi | no difference between number with preference for red and blue, but individuals were extremely flower constant. | Goulson & Cory (1993) |
| Gonepteryx rhamni | red/orange/yellow & blue/purple | Ilse (1928) |
| Vanessa polychloros | red | Ilse (1928) |
| Argynnis paphia | primary: orange/yellow secondary: blue/purple | Ilse (1928) |
| Macroglossum stellatarum | primary: 440nm secondary: 540nm | Kelber (1996) |
| Inachis io | yellow & blue | Ilse (1928) |
| Papilio machaon | blue/purple | Ilse (1928) |
| Papilio demoleus | blue/purple | Ilse & Vaidya (1956) |
| Papilio troilus | primary: blue secondary: orange | Swihart (1970) |
Butterflies have also been shown to learn to be efficient in their foraging by distinguishing rewarding and non-rewarding flowers, and learning where to probe in a particular species (Kandori & Ohsaki, 1996; Kandori & Ohsaki, 1998). They can also learn rapidly to feed from different coloured flowers if these are rewarding (Swihart & Swihart, 1970; Goulson and Cory, 1993; Weiss 1995, 1997; Kandori & Ohsaki, 1996; Kelber 1996; Lewis & Lipani, 1990), although increasing the number of species from which they have learnt to feed appears to reduce their efficiency in feeding from the flowers, possibly indicating memory constraints (Lewis, 1986). It might be expected that butterflies be able to learn which flowers in their particular environment give the best reward, as this may decrease the amount of time they have to spend foraging and therefore increase the amount of time they can spend searching for mates, laying eggs etc. The innate colour preferences may be explained as a way in which the individuals can distinguish flowers for feeding from a predominantly green or spectrally impure background (Klein, 1978; Menzel, 1979) when they first emerge. However, it is not easy to see why different species should have different innate colour preferences when they are not specialised to feed from flowers of a particular colour, so that the nutritional value of the flowers to the butterflies is not related to their colour. This may, therefore, be another case in which butterflies demonstrate a non-functional 'inherent bias' for a particular colour or pattern. In Chapter 2, the flower and mate colour preferences of the polymorphic butterfly Argynnis paphia were investigated, and it was found that the males had a preference for orange females (confirming the work of Magnus, 1958) which was also reflected in their preference for orange flowers (Ilse, 1928). In the case of Argynnis paphia it was not possible to determine whether this preference for orange had evolved for the purpose of mate recognition and simply had the added effect of causing an attraction for orange flowers, or whether the preference was due to an evolutionarily neutral colour bias which was then expressed in both flower and mate choice. Cook et al. (1994) demonstrated a preference in Papilio dardanus for true white morphs (hippocoonides) over yellow morphs (trimeni) over red/orange morphs (lamborni) (see Chapter 3 for an analysis of the colours). Since the white morphs are the most common this could be explained in terms of a rapid learning effect, whereby naďve males mate at random, and then subsequently mate more frequently with females of the same colour. It could also be explained by innate colour preferences, which could be expressed in both mate and flower choice.
The aims of this chapter are to determine the innate flower colour preferences of Papilio dardanus, and to investigate the degree of constancy it shows to colours, indicating the colour learning capabilities of the species. These can then be compared with the mate choice behaviour studied in Chapter 5.
As summarised in Table 4-1, different species of butterfly have been shown to have different colour preferences in their naďve choice of flower. Crane (1955) described 'tropical Papilionids and Pierids' as feeding predominantly from red flowers at her research station in Trinidad. However, experiments carried out to test the naďve preference of three species of the genus Papilio showed a primary preference for flowers in the blue area of the spectrum (Papilio machaon, Ilse, 1928; Papilio demoleus, Ilse & Vaidya 1956; Papilio troilus, Swihart, 1970), as did two out of the three experiments carried out on species of the genus Pieris (Pieris brassicae, Scherer & Kolb, 1987; Pieris rapae, Kandori & Ohsaki, 1996; equal red/blue preference in Pieris napi, Goulson & Cory, 1993). This suggests that these butterflies have an initial preference for the blue region of the spectrum which, at least in some geographic regions, may subsequently be altered by learning to allow the butterflies to exploit the most rewarding flowers.
The order of colour preferences found in mate choice in Papilio dardanus by Cook et al. (1994) could be due to an innate bias for certain colours or, since the ranking of morph preferences reflects the proportions of the different morphs, due to a learning effect in males. Some butterflies have been shown to demonstrate a high degree of flower constancy (Goulson & Cory, 1993; Kandori & Ohsaki, 1996) - returning again and again to the same colour. This may indicate that they can learn that a certain colour flower is profitable from a single visit, and this type of 'colour imprinting' could explain the mate choice preferences of Papilio dardanus.
The aim of this Experiment is to ascertain the initial flower colour preference of naďve Papilio dardanus butterflies, and to measure the degree of flower colour constancy shown by individuals in order to assess the rapid colour learning capabilities of the species.
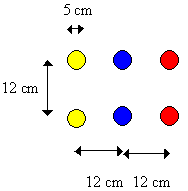
Individual Papilio dardanus of either sex which had not fed from flowers before were placed in a flight cage (see Appendix 3 for dimensions and further details) - up to 15 individuals were present in the cage at any one time. In the centre of the cage was placed a 2 x 3 array of artificial flowers (diameter 5cm, resembling poppies - see Figure 4-1) each containing sugar water at a concentration of approximately 1 part sugar to three parts water. The flowers were painted red (reflectance 640nm and above), blue (reflectance between 400-500nm with peak at 460nm) and yellow (reflectance above 580nm) (all enamel paint, Humbrol Ltd., Hull; reflectance spectra from the manufacturer), and placed in an array. None of the paints had any UV reflectance. It was decided not to include a true-white (achromatic) flower, which would represent the most favoured colour in the case of mate preference. This is because discrimination between different hues and different brightnesses of achromatic stimuli are carried out by different neural mechanisms, and butterflies show an almost linear relationship between the brightness and attractiveness of achromatic flower models (Swihart, 1971). Therefore only red and yellow were used in the Experiment, and compared with blue, which has been shown to be innately attractive to Papilionids (see Table 4-1). The array was placed across the width of the rectangular cage (East-West), with 12 cm between adjacent flower centres and adjacent flowers of each of the three pairs being the same colour (See Figure 4-2). Pairs of flowers were used in order to decrease the effect of any individual differences between the flowers which might affect the recorded colour choices of the butterflies.
As soon as an individual landed on a particular flower and extended its proboscis, the colour and position (North or South) of the flower on which it landed was recorded. The butterfly was allowed to feed, and the underside of its wings were marked with a permanent marker to allow subsequent visits by the same individual to be recorded. The length of the visit to the flower was also classified as 'brief' (under approximately 3 seconds), 'medium' (approximately 3-10 seconds), or 'long' (over 10 seconds). After five naďve butterflies had been recorded as making an initial colour choice, the order of the colours was changed to average out any spatial effects, and this was repeated after every batch of five initial choices. Subsequent visits by individuals were also recorded to allow an assessment of colour constancy.
Butterflies approached the flower array from all directions, but mainly from either the North or South as they flew up and down the length of the cage.
| South | North | Total | |
|---|---|---|---|
| Blue | 16 | 9 | 25 |
| Red | 0 | 1 | 1 |
| Yellow | 3 | 1 | 4 |
| Total | 19 | 11 | 30 |
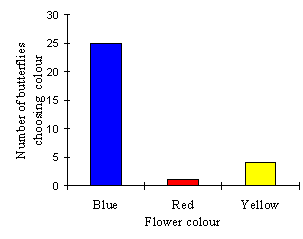

This data indicates that there is no significant preference for flowers at the North or South of the array (p=0.100, binomial test, n=30), but that there is a significant preference for blue over red and yellow flowers (p< 4x10-8, (c2= 34.2, n=30).
| South | North | Total | |
|---|---|---|---|
| Blue | 117 | 110 | 227 |
| Red | 4 | 1 | 5 |
| Yellow | 28 | 25 | 53 |
| Total | 149 | 136 | 285 |
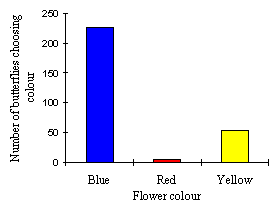

The data for all recorded flower choices shows a distribution which is not significantly different from this (p=0.135, n=285, Friedman Test).
A constancy index was used, where:
and:
These were calculated for each butterfly which had visited the flowers more than once and a mean value for each obtained:
constancy to 1st choice = 0.90 (standard deviation 0.21, n=30)
constancy to favourite colour = 0.927 (standard deviation 0.149, n=30)
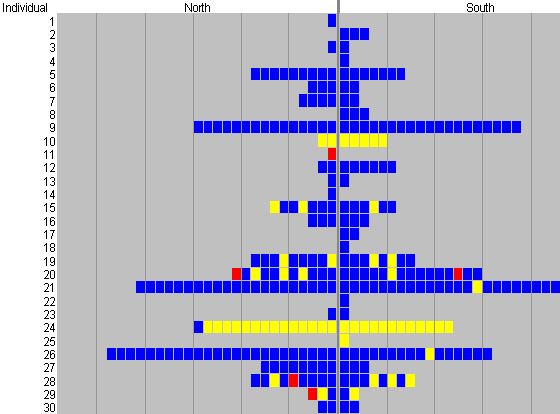
These figures are very similar, as only one butterfly (no. 19, see Figure 4-5) actually visited a flower first which was not subsequently visited the most often. In addition, only two individuals (nos. 10 & 24) had favourite colours which were not blue, but it is interesting that both of these showed very high constancy to this colour (100% in the case of butterfly 10, and 96% in the case of butterfly 24). This suggests that the high degree of constancy shown may be due to learning rather than innate preference (unless there is a degree of polymorphism in the innate preference).
Since butterflies were often observed to alternate between the North and South flower of the same colour, the number of visits to the same and the alternative position (either 'North' or 'South' regardless of colour) were counted:
| Number of visits to the same position | Number of visits to the alternative position |
|---|---|
| 83 | 162 |
This shows that the butterflies are visiting the flower at the alternative position on the array (despite their high colour constancy) significantly more often than returning to the same position (p=1.3x10-7, binomial test, n=245).
This Experiment demonstrates that Papilio dardanus show a significant preference for blue flowers over yellow and red. They are not significantly changing their behaviour on subsequent visits, and show no significant spatial preferences within the array. They show a high degree of constancy (90%) to a colour when the flowers provide a reward, and this colour is almost always the first to which the butterfly has made a visit - only one individual had a favourite colour which was not its first choice. It therefore appears that the butterflies are learning a profitable flower colour from their first visit, and subsequently visit the same colour 90% of the time.
It is interesting to plot a graph to show the length of the feed (classified as 'brief', 'medium' or 'long'), and to compare the overall proportions of the three feeding lengths (representing 'normal feeding') compared with the categorised length of the feed just before a butterfly chooses to feed off a new colour:
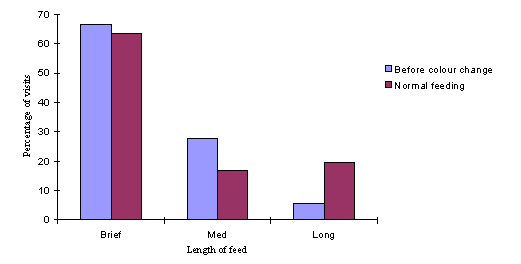
These data suggest that the butterflies tend to try a new colour when they have failed to feed successfully (indicated by a 'brief' visit) from their initial colour choice. This is backed up by the evidence from one butterfly (who up until that point had fed exclusively off yellow flowers) to an array of empty flowers. This data was gathered when the array was introduced to the flight cage full of butterflies at the start of an experimental session. The butterfly came directly down to the array before the flowers had been filled up with sugar water. Her choices in order were:
| Blue | Yellow | Red | |||
|---|---|---|---|---|---|
| South | North | South | North | South | North |
| 1 | |||||
| 2 | |||||
| 3 | |||||
| 4 | |||||
| 5 | |||||
| 6 | |||||
| 7 | |||||
| 8 | |||||
In this sequence the butterfly showed highly uncharacteristic colour swapping (and it is unusual that she starts the searching with a blue flower) - she had never been to a flower other than yellow, and when the flowers were filled with sugar water she fed almost exclusively off yellow once again (see individual 24 in Figure 4-5). This supports the idea that the butterflies are learning rapidly, from their first profitable feed, and will stick to the first colour from which they feed successfully, only trying other colours when they fail to find food from their original colour choice.
The butterflies also show a significant alternating behaviour between the pairs of flowers of the same colour (see Table 4-4). This presumably increases their chances of finding nectar in a natural situation, as they may have emptied one flower on their last visit and it may need time to refill. In addition, it was also noted that the butterflies appeared to try more colours when there were more individuals feeding - several of the cases of a butterfly choosing a colour other than its usual favourite occurred when other individuals were feeding at the array - and this is investigated in later experiments.
In Experiment 4-1, all the flowers had been painted with the same enamel paint (from Humbrol Ltd, Hull). To investigate whether or not the glossy painted surface was more or less attractive to the naďve butterflies than a more natural, matt, surface, a pilot Experiment was designed using unpainted artificial flowers, since even matt paint tended to give a gloss finish when used on the woven plastic surface of the artificial flowers. The artificial poppies used in Experiment 4-1 were originally red in colour, before being painted, and had a naturally matt texture. The original red colour was likely, however, to have a slightly different spectrum from the red of the enamel paint, and comparing the two directly in a choice test might cause the butterflies to distinguish between the two colours rather than show a preference for the texture. Since red was significantly less preferred than blue and yellow in Experiment 4-1, it was decided to use the same artificial flowers as before, but to leave one pair unpainted red in order to compare the number of visits to the unpainted red flowers with the number in the previous Experiment to the painted red flowers. It was hoped that this method might be more successful at detecting a texture preference than a direct comparison between unpainted and painted red flowers.
The aim of this Experiment is to determine whether or not naďve Papilio dardanus show a significant preference for red flowers over blue and yellow when the red flowers are left unpainted, with a matt texture, whilst the blue and yellow flowers have a gloss paint finish.
Individual Papilio dardanus of either sex which had not fed from flowers before were placed in the flight cage, with up to 20 individuals in the flight cage at any one time. In the centre of the cage was placed a 2 x 3 array of artificial flowers (diameter 5cm, resembling poppies), as in Figure 4-2. Two artificial flowers were left unpainted, with a matt red surface, and the others were painted blue and yellow (enamel paint, Humbrol Ltd., Hull), and placed in the array, with 12 cm between adjacent flower centres, adjacent flowers of each of the three pairs being the same colour, as in Experiment 4-1 (see Figure 4-2). As before, pairs of flowers were used to decrease any effects introduced by individual differences between the flowers. Since in this Experiment only the initial flower choices of the naďve butterflies was being recorded, the flowers were left empty, with no sugar water in them. As soon as an individual landed on and tried to feed from a particular flower, it was removed from the cage, and the choice recorded to determine the initial preferences of the butterflies. After every ten data points were recorded, the order of the colours was changed to average out any spatial effects from the positions of the flowers.
| Number choosing blue (gloss) | Number choosing yellow (gloss) | Number choosing red (matt) |
|---|---|---|
| 1 | 2 | 27 |
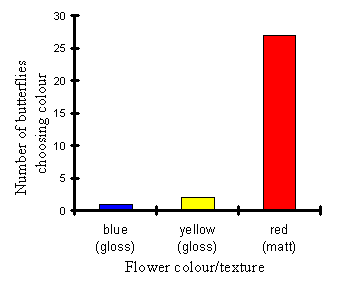
The results indicate that the Papilio dardanus now have a strongly significant preference for the matt red flowers over the gloss blue and yellow (p < 4x10-10, c2 = 43.4, n=30). This is a very big shift in behaviour when compared with the results of Experiment 4-1 where naďve butterflies showed a significant preference for blue flowers when all were painted with colours having the same gloss finish.

The results of this Experiment indicate that the butterflies are not choosing between the flowers simply on the basis of their colours, and that there is another factor determining their choice which appears to be more important to them and can shift their colour preferences dramatically. Unfortunately, there were differences between the painted and unpainted red flowers other than texture in this pilot Experiment, such as the darkness of the central area (see Figure 4-8), which had been painted over in Experiment 4-1. A further Experiment was therefore planned to investigate the differences between the unpainted red flowers and the painted red flowers which might be affecting the choices of the butterflies.
The previous experiments showed that the butterflies significantly preferred unpainted red flowers over painted blue and yellow flowers, but significantly preferred blue flowers over red and yellow when all three colours were painted using the same gloss enamel paint. Therefore the choices of the butterflies appear to be influenced by a cue other than colour. Two differences between the painted and unpainted flowers were immediately obvious - firstly the unpainted flowers were made of a plastic fabric which had a matt finish rather than the uniform gloss finish of the painted flowers, and secondly they had black central markings, which could act as honey guides, which had been painted over in the painted flowers. It seemed likely that either of these two factors could be important (Kandori & Ohsaki have demonstrated the importance of honey guides, 1998), and therefore an Experiment was designed to test whether either of these could be influencing the butterflies' choices.
The aim of this Experiment is to ascertain whether or not the butterflies show a preference for matt flowers over gloss flowers, and whether or not they are attracted to flowers with a dark central area over those without one.
Naďve Papilio dardanus of either sex were placed in the flight cage, with up to 20 individuals in the cage at any one time. In the centre of the cage, as in Experiments 4-1 and 4-2, was placed an array of the same artificial flowers. The array in this Experiment was made up of only four flowers, although the distances between the centres of the flowers were still 12cm (see Figure 4-9) - two were painted red (using the Humbrol enamel paint), but on one of these black central markings were made with a permanent marker pen similar to the markings on the unpainted red flowers. Two were painted blue - one gloss as before, but the other was made matt by covering the petals with cloth and painting the cloth with the same enamel paint (from Humbrol). This gave a matt finish to the flower.
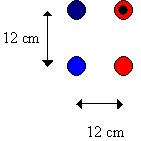
The flowers were filled with sugar water (approximately three parts water to one part sugar), and naďve Papilio dardanus were allowed to feed from them freely as in Experiment 4-1. When a butterfly landed on the array for the first time, its initial choice of flower was recorded, and the underside of its wings was marked with a permanent marker as in Experiment 4-1 without disturbing it from feeding. This allowed the butterflies to be recognised individually, so that subsequent visits to the flowers by the same individuals could be recorded. The flower array was rotated 180o after 15 data points had been recorded to remove any spatial effects from the results, but the two gloss flowers were always adjacent, as were the matt and the black centred flowers.
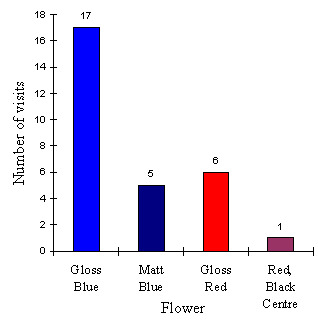
The results of the first choice of butterflies is shown in Table 4-7 and Figure 4-10.
| Gloss Blue | Matt Blue | Plain Red | Red, Black Centre | |
|---|---|---|---|---|
| No. 1st Visits | 17 | 5 | 6 | 1 |
The distribution of subsequent choices is shown in Figure 4-11:
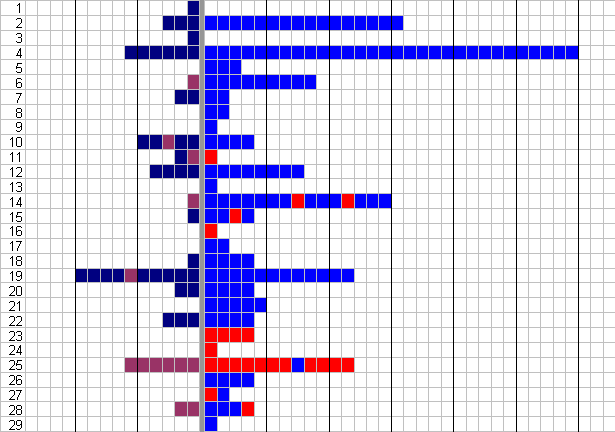
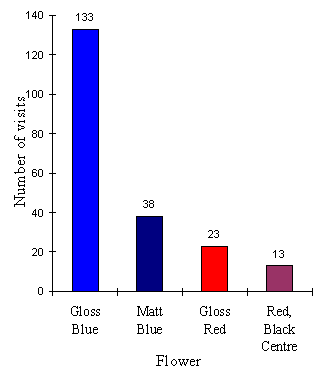
The results show that the butterflies display a significant preference for gloss blue flowers over any of the other options on their first choice of flower (p<0.0003, c2 =19.4, n=29, ), and an even greater bias for gloss blue on their subsequent choices, although this may well be due to the fact that the fabric of the matt flower soaked up sugar water and thus the flower was less rewarding. There is no significant difference between the overall choices of red flowers and red flowers with a black centre (p=0.066, binomial test, n=36). Calculations of the butterflies' constancy to a particular flower can be calculated as in Experiment 4-1:
Constancy to 1st flower chosen = 0.660 (standard deviation = 0.28, n=29)
Constancy to favourite flower = 0.733 (standard deviation = 0.22, n=29)
These figures are lower than the constancies recorded in Experiment 4-1, but this is probably due to some individuals failing to distinguish between the matt and gloss blue flowers, and between the red flower and red flower with a black centre. If the choice is reduced to simply red or blue, the constancies recorded are:
Constancy to 1st flower chosen = 0.871 (standard deviation = 0.234, n=29)
Constancy to favourite flower = 0.908 (standard deviation = 0.158, n=29)
These are close to the values of 0.90 and 0.927 recorded in Experiment 4-1.
These results suggest that neither the matt texture nor the presence of black central markings in the flowers are significantly attractive to Papilio dardanus. In fact the gloss finish was significantly preferred to the matt finish. The butterflies remain highly constant to a particular flower, but many appear not to discriminate between the matt and gloss and the red flower and red flower with black centre, simply remaining 90% constant to a particular colour. It therefore appears that the butterflies are preferring a gloss finish to the flowers, and that they do not significantly prefer the red flowers with the black centre to the plain red flowers (in fact they show an insignificant preference for the flower without the black centre). The chemical constituents of the paint itself would not seem to be particularly repellent, as the blue-painted flowers used in Experiment 4-1 were the most freshly painted (only hours before the first experimental session), and were strongly preferred over the red and yellow flowers which had been painted some months previously. Many of the chemicals present in the flowers would also not be detectable to the butterflies until they had landed, when tarsal receptors may be used. It is possible that there is some feature of the artificially matt flower in Experiment 4-3 which is unattractive to them and therefore biasing the results. Unfortunately further experiments to investigate this phenomenon were not possible due to time constraints, and the fact that further artificial flowers of the same design were unobtainable. The preference for the unpainted red flowers in Experiment 4-2 therefore remains unexplained.
Whilst Experiments 4-1 and 4-3 were being carried out, it was noticed that the Papilio dardanus tended to feed more often, and from a wider range of colours when other individuals were feeding from the array. In fact some of the instances where an individual fed off a flower which was not its 'favourite' colour in Experiment 4-1 coincided with other butterflies being present on the array (e.g. individual 29's final three visits in Figure 4-5). It thus seemed that although Papilio dardanus had an initial preference for blue, they could also learn from other individuals to feed from other colours. This is another aspect of feeding behaviour which appears not to have been investigated before, and could be a very interesting way for naďve butterflies to increase their feeding efficiency rapidly.
An Experiment was therefore planned to test whether the presence of another individual feeding on the array would influence the colour choices of other individuals. In order to simulate the presence of an individual feeding at the array, a dead butterfly (with its wings open) was used, pinned on to one of the flowers. This allowed complete control of the appearance of the 'teacher' individual.
This Experiment aims to determine whether the apparent presence of another individual feeding affects the colour choice of naďve Papilio dardanus.
Since an Experiment had already been carried out on the initial colour preferences of Papilio dardanus, without the influence of other individuals feeding (Experiment 4-2, in which individuals were removed as soon as they had alighted on a flower in order to feed), the results from Experiment 4-2 could be used as the control for an Experiment on the influence of another individual at the array. Therefore, the same array as had been used in Experiment 4-2 (with unpainted red flowers versus painted blue and yellow flowers) was used in this Experiment.
Naďve Papilio dardanus of either sex were placed in the flight cage, with up to 20 individuals in the cage at one time. The array of flowers used in Experiment 4-2 (see Figure 4-2) was placed in the centre of the cage as before. A dead male Papilio dardanus, with wings open, was pinned onto a flower of each colour in turn (being moved after every 5 data points was recorded) so as to be obvious from every approach angle but not obscuring the flower. When a butterfly landed on a flower and extended its proboscis, the flower colour and position was recorded, as was the sex of the butterfly concerned. It was then removed immediately so as not to influence the choice of any of the other butterflies in the cage.
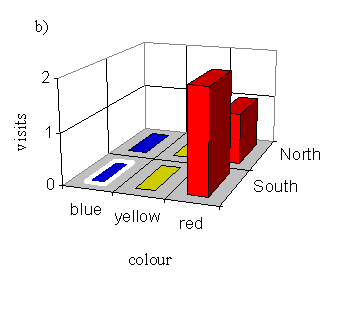
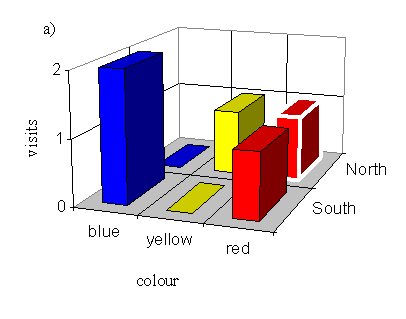
With a male pinned onto a blue flower (order blue, yellow, red):
| Number choosing blue | Number choosing yellow | Number choosing red |
|---|---|---|
| 2 | 1 | 7 |

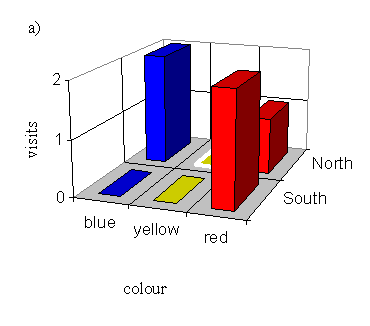
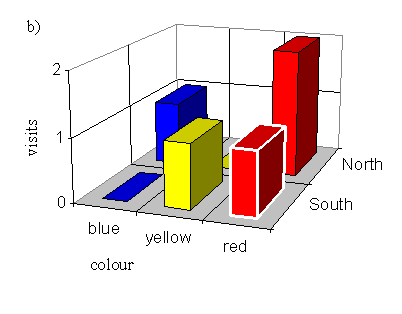
With a male pinned onto a yellow flower (order blue, yellow, red):
| Number choosing blue | Number choosing yellow | Number choosing red |
|---|---|---|
| 4 | 0 | 6 |
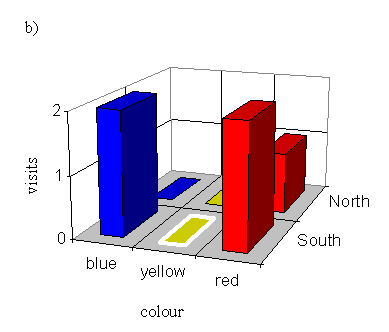
With a male pinned onto a red flower (order blue, yellow, red):
| Number choosing blue | Number choosing yellow | Number choosing red |
|---|---|---|
| 3 | 2 | 5 |
Taking the data as a whole, the totals are:
| Number choosing blue | Number choosing yellow | Number choosing red |
|---|---|---|
| 9 | 3 | 18 |
Comparing the results of this Experiment, shown in Table 4-11, with those of Experiment 4-2, shown in Table 4-6, indicates that the pinning of a dead male onto various flowers has made a significant difference to the initial choices of the butterflies (using Table 4-6 to generate the probabilities of choosing red or other colour, and then using these to perform a binomial test on the results in Table 4-11: p = 0.000015, n=30). The yellow and blue flowers are now visited significantly more often.
Splitting the colour choices into those gathered from males and those from females, the results were:
| Number choosing blue | Number choosing yellow | Number choosing red | |
|---|---|---|---|
| Males | 7 | 2 | 11 |
| Females | 2 | 1 | 7 |
This data set is difficult to analyse statistically (as the numbers of visits to yellow and blue flowers are so small), but it is possible to see that the two sexes rank the colours in the same order, and appear to show very similar behaviour.
Although the Experiment shows that pinning a dead butterfly onto a flower does alter the choice of other butterflies, they do not simply seem to choose flowers of the same colour as that onto which the dead butterfly is pinned. In fact, when the butterfly is pinned on a blue flower (see Table 4-8), only two individuals chose blue (as opposed to four and three when the butterfly was pinned onto yellow and red respectively - see Table 4-9 and Table 4-10). When the butterfly was pinned on a yellow flower (see Table 4-9), yellow was never chosen (as it was once and twice when the butterfly was pinned onto blue and red respectively - see Table 4-8 and Table 4-10). When the butterfly was pinned on a red flower (see Table 4-10), red was chosen only five times (rather than seven or six times when the butterfly was pinned onto blue and yellow respectively - see Table 4-8 and Table 4-9). The possibility that the butterflies are avoiding the flowers on which the dead one is pinned is ruled out by the fact that out of the seven occasions when the butterflies went to the same colour flower as the one on which a butterfly was pinned, three were visits to the same actual flower as the one on which the individual was pinned, and four were to the other one of the same colour.
On the basis of the data shown in Figure 4-13, Figure 4-14, and Figure 4-15 it is possible that the butterflies are choosing a neighbouring flower to the one on which an individual appears to be feeding. When the butterfly was pinned onto the North blue flower, this effect is clear (although the preference for red is still evident) - when the butterfly was pinned onto the South blue there is not enough data to show any effect. When the butterfly was pinned onto a yellow flower, the adjacent blue flower appears to get more visits, although there is oddly no visit at all to either yellow flower. When the butterfly was pinned on a red flower, the effect on the visits to the adjacent yellow flower seems to follow the same pattern. However, the data sets are very small as this Experiment was designed to test for a preference in colour rather than spatial position, and so another Experiment was designed in order to test the new hypothesis that the butterflies are attracted to a flower which is spatially close to the one on which another individual appears to be feeding.
Experiment 4-4 indicated that the presence of another individual (in the form of a dead male pinned onto a flower) affected the flower choice of other Papilio dardanus. The butterflies did not appear to be choosing flowers of the same colour as the one on which the male was pinned, but there was some evidence that they were choosing flowers which were spatially close to that on to which another individual had apparently landed.
'Imitation' has been reported in many vertebrates, but the terminology has been very confused (for reviews, see Galef, 1988 and Whiten & Ham, 1992). Many terms have been used for the type of behaviour which appears to be present in Papilio dardanus. Galef (1988) suggests 'social enhancement' to refer to the "simple presence of others, presence of behaving others, or presence of residual cues emitted by others .... [which] enhance performance of responses already in an individual's repertoire" (p.13). Within this umbrella term, the butterflies would be showing 'local enhancement' as defined by Thorpe (1963): "apparent imitation resulting from directing the animal's attention to a particular object or to a particular part of the environment" (p.134), or Miller & Dollard's (1941) 'matched dependent behaviour' in which individuals match their own behaviour to that of other individuals. As Galef (p.19) says, in behaviour of this type, "once the leader (the discriminatory stimulus) departs, those aspects of the follower's behaviour dependent on the presence of the leader are lost. For the pattern of behaviour initiated by the leader to become part of the behavioural repertoire of the follower, independent of the leader, the pattern of behaviour must come under the control of stimuli not dependent on the presence of the leader." So, in this case, by following the feeding behaviour of another butterfly and being rewarded, an individual could increase its repertoire of feeding behaviour. From Experiment 4-4 it appears that this is likely to be through spatial rather than colour learning. This Experiment is designed to test this, by removing the colour variation between the flowers and by increasing the size of the array to allow greater sensitivity to spatial positioning.
The aim of this Experiment is to determine whether or not Papilio dardanus choose flowers spatially close to those from which another individual appears to be feeding.
As was mentioned in the discussion in Experiment 4-3, the artificial flowers used in Experiments 4-1, 4-2, 4-3, and 4-4 became no longer available. Therefore, to produce a suitably large array of identical flowers, another design of artificial flower had to be used. Those which were most similar to (but slightly larger than) the artificial poppies used previously were chosen. The flowers were also placed closer together in order to fit a linear array across the width of the flight cage.
Naďve butterflies were introduced to the flight cage (with up to twenty individuals present at one time) in the centre of which was an array of nine red artificial flowers resembling poppies, with an average diameter of 8cm, and with an average separation of 9.5cm from the centre of one to the centre of the next in line. The array consisted of nine flowers in a row (see Figure 4-16)
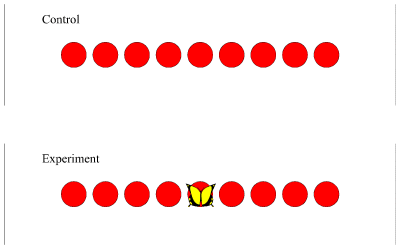
Since only the initial choice of the butterflies was being recorded, none of the flowers contained sugar water. In the control Experiment, the naďve butterflies were allowed to visit the flowers, and as soon as an individual landed on a flower and extended its proboscis, the butterfly was removed, and the position of the flower on which it landed was recorded. For the experimental condition, a dead male Papilio dardanus was pinned onto the central flower, making sure that it was obvious from all angles, and that it did not obscure the flower onto which it was pinned. Visits by naďve butterflies were recorded as in the control Experiment, and each butterfly was immediately removed as soon as it had landed on a flower and extended its proboscis so as not to affect the choice of other butterflies. After 15 visits had been recorded under each condition, the flower array was rotated by 180o (so that the flower which had been number 1 as labelled from west to east became number 9).
| Flower | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
| Control | 2 | 5 | 3 | 5 | 2 | 7 | 2 | 1 | 3 |
| Exptl. | 2 | 2 | 4 | 3 | 11 | 3 | 2 | 1 | 2 |
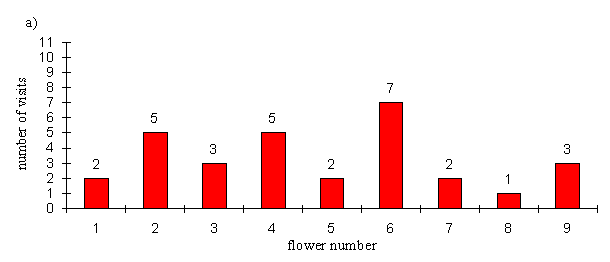
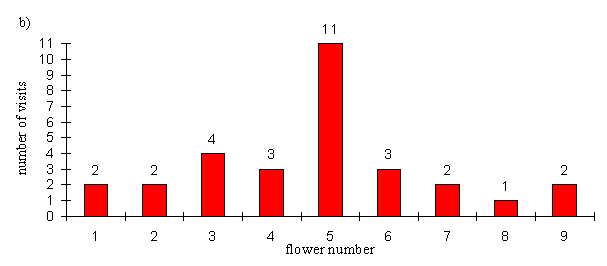
Under control conditions, the preferences of the butterflies were not significantly different from random behaviour (p=0.384, binomial test, n=30), whilst in the second they were significantly different (p=0.0021, binomial test, n=30). Of course the probability of getting 11 or more visits to the precise flower on which the other butterfly was pinned is even less. The visits to the other flowers in the experimental array are not significantly different from random (p=0.711, binomial test, n=19). So the butterflies appear to be choosing flowers at random when they are in an straight line, but when another individual is feeding on a flower they show a significant preference for that flower. It is interesting to note, however, that the preference for neighbouring flowers, seen in Experiment 4-4, appears not to be present. This may be related to the size of the flowers used, as these flowers were bigger than those used in Experiment 4-4. Their large size may indicate that there is likely to be enough nectar for two individuals to feed at the same time, or their size may simply make it easier for this to happen.
An analysis of the sexes of the butterflies involved (see Table 4-14), indicates that both sexes show the same trend. The behaviour of either sex in the control Experiment is not significantly different from random (binomial tests, p=0.69 for males, p=0.18 for females, n=15 in both cases). In the control Experiment, the females' preference for the central flower is significant (binomial test, p=0.001, n=16), and approaching significance for the males (binomial test, p=0.06, n=14). Given the relatively small data sets in each case, it seems safe to conclude that there is not a significant difference between the behaviours of the two sexes.
| Flower number | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Control | male | 0 | 4 | 2 | 3 | 1 | 2 | 1 | 0 | 2 |
| female | 2 | 1 | 1 | 2 | 1 | 5 | 1 | 1 | 1 | |
| Experimental | male | 1 | 0 | 2 | 3 | 4 | 1 | 2 | 0 | 1 |
| female | 1 | 2 | 2 | 0 | 7 | 2 | 0 | 1 | 1 | |
The presence of another individual feeding at an array of flowers appears to influence the feeding behaviour of other individuals of Papilio dardanus. The butterflies appear to match the behaviour of the individual already feeding, having a tendency to land on the same flower without showing any attraction towards the other individual as a mate or rival. It is not simply that the butterflies are being attracted by the sight of others to the feeding patch in general (as would be expected in most kinds of gregarious behaviour), but they are actually noting the exact feeding position of the other individual and copying that behaviour. From these experiments it is not possible to determine whether this has any effect on the butterflies' subsequent behaviour - i.e. whether they can be said to have learnt from the behaviour. However, given the rapid learning capabilities of butterflies as shown by Lewis & Lipani (1990) and others, it would seem quite possible that the individuals could learn to feed from certain flowers if rewarded when copying feeding behaviour seen in other individuals. This capability could allow them to learn to identify rewarding flowers more quickly, and possibly also to learn how to find nectar within them.
This behaviour may not be true 'imitation', as it has not been shown that the butterflies have actually copied a behaviour and incorporated it into their behavioural repertoire, although they can be said to be showing 'social enhancement' (Galef, 1988). This type of behaviour has been shown once in invertebrates before - Fiorito & Scotto (1992) showed that Octopus vulgaris will attack coloured balls having first seen a demonstrator do it in an adjacent tank. Although the example of Papilio dardanus feeding is not as impressive as the imitative behaviour of the Octopus, this appears to be the first demonstration of social facilitation in insects. In order to determine whether or not the butterflies are showing true 'imitation' behaviour (Whiten & Ham, 1992) it would be necessary to design an Experiment which would differentiate this from simply having their attention drawn to a food source. This would be difficult since firstly it is not possible to show that the butterflies are actually copying a behaviour pattern - the feeding reaction is innate and is carried out in the absence of other individuals - and secondly is not easy to show whether or not the choice of flower has been 'learnt' through the behaviour matching, as a subsequent test of the preferences of a butterfly which appeared to imitate on a first trial (in the presence of the other individual) would be greatly affected by the presence or absence of a reward on that first trial (an effect which would be difficult to remove).
The experiments in this chapter demonstrate that Papilio dardanus may have a relatively sophisticated feeding technique. They show a strong initial flower colour preference (preferring blue over yellow and red). This preference compares well with the experiments performed by Ilse (1928), Ilse & Vaidya (1956) and Swihart (1970) on other Papilio species which all demonstrated a primary preference for blue flowers. Comparing the preference with the ERG carried out in Experiment 3-5 it can be seen that the red paint has a reflectance which will barely be visible to the butterflies (longer than 640nm). The yellow (longer than 580nm) and blue (400-500nm) however, should both be clearly visible, exciting the pigments with peaks at 575nm (which would make the 'yellow' pigment appear red-shifted compared with our perception of it, due to the lack of green and UV reflectance) and 475nm (which would appear more green-shifted to them due to the lack of UV reflectance) respectively. Both these peaks are of a similar size, so the overwhelming preference for blue does not appear to be explained by the visual sensitivities of the butterfly. The preference, therefore (like the preference for orange found in Argynnis paphia in Chapter 2), must be due to a bias in higher processing. The fact that Crane (1955) observed tropical Papilionids and Pierids feeding mainly from red flowers in the field suggests that this initial preference for blue flowers is not adaptive. The preference for unpainted red flowers remains so far unexplained, but indicates that there is a factor other than colour which governs the innate preferences of the butterflies.
Experiment 4-1 demonstrated a high degree of colour constancy in the butterflies when the flowers offered a sugar water reward. It also presented a little evidence that if this reward was lacking, the butterflies were less constant, and tried other flowers. Experiments 4-4 and 4-5 also revealed that the butterflies could use social cues to improve their feeding success, trying a wider range of colours in Experiment 4-4 when another individual was present. This seemed to be due to butterflies feeding from flowers which were spatially close to that on which another individual appeared to be feeding regardless of the colour of the flower. Experiment 4-5 confirmed that the butterflies were indeed influenced by the presence of another individual feeding, but found that they were significantly more likely to feed off the same flower as another individual, rather than the neighbouring flower. The effect is likely still to be to increase the repertoire of colours from which an individual feeds, and possibly therefore the feeding efficiency, although Lewis (1986) found that increasing the number of species of flower from which an individual learnt to forage actually decreased their overall efficiency. This social effect is interesting, as it appears not to have been demonstrated before.
Cook et al. (1994) found a preference in male Papilio dardanus for yellow females over red females. In Experiment 4-1 there appeared also to be a preference for yellow over red (although the overwhelming preference for blue overshadowed this and made the two difficult to compare statistically), but the results of the ERG in Experiment 3-5 made it likely that this colour preference in flower choice was simply due to the visual sensitivities of the butterflies. Since Papilio dardanus shows a strong preference for blue flowers, it would be very interesting to test whether this colour preference could be present in their mate choice. Blue is a relatively unusual colour in nature as there are few blue pigments (although butterflies can use structural colours to produce blues (see Chapter 3)), but it is interesting to note that Papilio phorcas, the sister species of Papilio dardanus (Clarke et al., 1991;Vane Wright & Smith, 1991; Vane Wright et al., in press) does possess a blue pigment, phorcabilin (Choussy & Barbier, 1975) which it uses in association with yellow pigment to form its green coloration (Vane Wright et al., in press). This is discussed further in Chapter 5.
I am very grateful to Humbrol Ltd. for supplying the reflectance spectra for their enamel paints, and to Carol & Tony Alden for the artificial flowers. I would also like to thank Phil Taylor for his help with the flight cage, and John Calvert at Stratford Butterfly Farm for providing the butterflies.
Choussy, M. & Barbier, M. 1975. Ptérobiline et Néoptérobilines, réactivité et structures. Helvetica Chimica Acta 58, 2651-2661.
Darwin, C. 1877. The effects of cross- and self-fertilization in the vegetable kingdom. Murray, London.
Clarke, C.A., Gordon, I.J., Vane-Wright, R.I., Smith, C.R. 1991. Phylogenetic relationships of three African swallowtail butterflies, Papilio dardanus, P. phorcas and P. constantinus: new data from hybrids (Lepidoptera: Papilionidae). Systematic Entomology 16, 257-273.
Cook, S.E., Vernon, J.G., Bateson, M., Guilford, T. 1994. Mate choice in the polymorphic African swallowtail butterfly, Papilio dardanus: male-like females may avoid sexual harassment. Anim. Behav. 47, 389-397.
Crane, J. 1955. Imaginal behavior of a Trinidad butterfly, Heliconius erato hydara Hewitson, with special reference to the social use of color. Zoologica (N.Y.) 40, 167-196.
Fiorito, G. & Scotto, P. 1992. Observational learning in Octopus vulgaris. Science 256, 545.
Frisch, K. Von. 1914. Der Farbensinn und Orientierung der Biene. Zool. J. Physiol. 37, 1-238.
Galef, B.G. 1988. Imitation in animals: history, definition, and interpretation of data from the psychological laboratory. In: Social learning. Psychological and biological perspectives (Ed. Zentall, T. & Galef, B.). Hillsdale, New Jersey: Lawrence Erlbaum. 3-28.
Goulson, D. & Cory, J.S. 1993. Flower constancy and learning in foraging preferences of the green-veined white butterfly Pieris napi. Ecol. Entomol. 18, 315-320.
Ilse, D. 1928. Über den Farbensinn der Tagfalter. Z. f. vergl. Physiologie Bd. 8, 658-692.
Ilse, D. & Vaidya, V.G. 1956. Spontaneous feeding response to colours in Papilio demoleus L.. Proc. Indian Acad. Sci., Sect. B 43, 23-31.
Kandori, I. & Ohsaki, N. 1996. The learning abilities of the White Cabbage butterfly, Pieris rapae, foraging for flowers. Res. Popul. Ecol. 38(1), 111-117.
Kandori, I. & Ohsaki, N. 1998. Effect of experience on foraging behavior towards artificial nectar guide in the cabbage butterfly, Pieris rapae crucivora (Lepidoptera: Pieridae). Appl. Ent. and Zoo. 33(1), 35-42.
Kelber, A. 1996. Colour learning in the hawkmoth Macroglossum stellatarum. J. Expt. Biol. 199, 1127-1131.
Klein, R.M. 1978. Plants and near-ultraviolet radiation. Bot. Rev. 44, 1-127.
Lewis, A.C. 1986. Memory constraints and flower choice in Pieris rapae. Science 232, 863-865.
Lewis, A.C. & Lipani, G.A. 1990. Learning and flower use in butterflies: hypotheses from honey bees. In Insect-Plant Interactions, Vol. II, ed. Bernays, E.A., CRC Press, Boca Raton, Florida. 95-110.
Magnus, D. 1958. Experimentelle Untersuchungen zur Bionomie und Ethologie des Kaisermantels. Z. Tierpsychol. 15, 397-426.
Menzel, R. 1979. Spectral sensitivity and colour vision in invertebrates. In Handbook of Sensory Physiology Vol. VII/6A "Comparative physiology and evolution of vision in invertebrates" ed. Autrum, H., 503-580. Springer, Berlin.
Miller, N.E. & Dollard, J. 1941. Social learning and imitation, New Haven: Yale University Press.
Scherer, C. & Kolb, G. 1987. The influence of color stimuli on visually controlled behaviour in Aglais urticae L. and Pararge aegeria L. (Lepidoptera). J. Comp. Physiol. A 161, 891-898.
Swihart, C.A. & Swihart, S.L. 1970. Colour selection and learned feeding preferences in the butterfly, Heliconius charitonius L.. Anim. Behav. 18, 60-64.
Swihart, S.L. 1970. The neural basis of colour vision in the butterfly Papilio troilus. J. Insect Physiol. 16, 1623-1636.
Swihart, C.A. 1971. Colour discrimination by the butterfly, Heliconius charitonius L.. Anim Behav. 19, 156-164.
Thorpe, W.H. 1963. Learning and instinct in animals. 2nd Edition. London: Methuen.
Vane Wright, R.I. & Smith, C.R. 1991. Phylogenetic relationships of three African swallowtail butterflies, Papilio dardanus, P. phorcas and P. constantinus: a cladistic analysis (Lepidoptera: Papilionidae). Systematic Entomology 16, 275-291.
Vane Wright, R.I., Raheem, D.C., Cieslak, A., Vogler, A. In press. Evolution of the mimetic African swallowtail butterfly Papilio dardanus: molecular data confirm relationships with P. phorcas and P. constantinus. Biol. J. Linn. Soc.
Weiss, M.R. 1995. Associative colour learning in a nymphalid butterfly. Ecol. Entomol. 20, 298-301.
Weiss, M.R. 1997. Innate colour preferences and flexible colour learning in the pipevine swallowtail. Anim. Behav. 53, 1043-1052.
Whiten, A. & Ham, R. 1992. On the nature and evolution of imitation in the animal kingdom: reappraisal of a century of research. Advances in the Study of Behaviour 21, 239-283.
|
Please cite this thesis as: Freeman, ALJ; 1998; D.Phil thesis, Oxford University. |
E-mail to Alexandra Freeman Back to Table of Contents Back to Home Page |