
Figure 6-1 A gynandromorph of Papilio dardanus showing the difference in morphology of males and females, used to predict palatability in Experiment 6-6
The theory of mimicry requires that some individuals are less profitable to predators than others. The actual palatability of a species, however, is not easy to ascertain as it depends on many factors. Firstly, different predators may have different tastes. For example, some bird species (Pheucticus melanocephalus and Icterus galbula) and mice (Peromyscus melanotis) have been shown to eat overwintering Monarch butterflies (Danaus plexippus), being resistant to their poisons (Fink & Brower, 1981, Glendinning & Brower, 1990). The palatability of a particular prey item to the predator also depends on the psychological state of the predator - for example how hungry it is (Swynnerton, published posthumously by Carpenter, 1941). Secondly, individuals within a species may have differing palatabilities, for example Monarch butterflies which have fed on milkweed species, such as Asclepias curassavica, which contain cardiac glycosides are able to sequester the chemicals (calactin, calotropin and calotoxin), whilst those which have fed on species lacking these emetic chemicals, such as Gonolobus rostratus are completely palatable to bird predators (Brower, 1969). This results in a 'palatability spectrum' (Brower et al., 1968) of individuals within and between species for a particular predator. The point at which the predator distinguishes 'unpalatable' from 'palatable' depends on the state of that individual.
Given these complications, experiments to test the relative palatabilities of different prey items are difficult to plan and carry out. The simple chemical identification of a compound in a prey item which can be shown to be unpalatable to a major predator may not be evidence that the predator would avoid the prey item under natural conditions. The most naturalistic experiments involve releasing captured butterflies to wild birds feeding from a perch (e.g. Swynnerton, 1915, 1919; Carpenter, 1941), but this only tests the perceived palatability of the prey to the predator, which may be mistaking mimetic species for their models, and also needs to be carried out throughout the year to measure the average palatability (e.g. Srygley & Kingsolver, 1998). In order to overcome this problem, the coloration of the prey could be disguised, and naďve predators used.
Papilio dardanus is a classic example of Batesian mimicry. The palatable females mimic unpalatable species in order to gain protection from predators. However, in none of the experiments by Swynnerton (Carpenter 1941) in which he released wild-caught butterflies to wild birds (mostly the bee eater Merops merops) was a male Papilio dardanus eaten by the birds, although two were caught and immediately released. This suggests that the butterflies may have at least appeared to be distasteful to the birds. In addition, the wings of the species smell strongly of pyrazines, chemicals thought to act as a chemical signal of distastefulness in combination with bright warning colours, such as those found in Papilio dardanus (Rowe & Guilford, 1996), and which are difficult to isolate and identify chemically, requiring gas chromatography and mass spectrometry (Rothschild et al., 1984).
There is much debate about the reason why a male-like andromorph is found in some populations of Papilio dardanus (such as the tailed yellow females in races antinorii, meriones, and humbloti, and the tailless morph trimeni in race tibullus). Many argue that the male-like female is ancestral (Poulton, 1924; Ford, 1936; Clarke & Sheppard, 1963; Turner, 1963; O'Donald & Barrett, 1973; Clarke et al., 1985), but do not explain how the morph is maintained in a population which includes Batesian mimics which must be at an advantage in avoiding predation. Where the race is entirely monomorphic, such as races meriones and humbloti, it can be argued that the mimetic morphs have simply not evolved (possibly due to a lower degree of predation as these morphs are confined to islands where bird fauna is impoverished) (Trimen, 1868). Others argue that the male-like females may be derived (van Bemmelen, 1922; Bernardi, 1963; Vane-Wright & Smith, 1991, Vane-Wright et al., in press), possibly mimicking the males in order to avoid sexual harassment (Cook et al., 1994) although this latter hypothesis seems less likely in the light of the observations and results of Chapter 5. The fact that their genes are dominant over those of most other female morphs (excluding lamborni) in the populations in which they exist suggests that they may have a more recent origin (Sheppard, 1967), although, as discussed in the conclusions to Chapter 3, the tendency for new genes to become dominant has been shown mathematically to be true only under certain conditions (Charlesworth & Charlesworth, 1976). In Chapter 5 it was shown that the males appear to have no morph preferences when mating for the first time but subsequently show a preference for the morph with which they first mated, and therefore that the andromorphic females were unlikely to be at any advantage through sexual selection. Whether they are merely the ancestral morph, maintained in the population, or whether they are a recently evolved male-mimic, there must be some selective pressure resulting in them being present in the polymorphic, mimetic populations.
The presence of a pyrazine odour on the wings of Papilio dardanus is likely to reinforce the warning effect of the coloration on the wings of the mimetic morphs (Rowe & Guilford, 1996), but all individuals carry the same odour, and so the bright yellow of the males and andromorphs may have the same warning effect in association with the pyrazine odour. Rowe & Guilford also found, however, that the signal loses its significance if the warning odour and coloration is not backed up by unpalatability. If the species is in fact slightly unpalatable then the yellow coloration and pyrazine odour would act as an honest warning. The effect seen by Swynnerton (Carpenter, 1941) of two birds releasing male Papilio dardanus once they had been caught does accord with the hypothesis that they were deterred from eating the butterflies because of the pyrazine odour they emitted when caught. It is possible therefore that the species is not, in fact, completely palatable to its major predators. The existence of a palatability spectrum (Brower et al., 1968) means that many cases of mimicry might be expected to fall between the standard definitions of Batesian and Mullerian mimicry, and the population dynamics and evolutionary pressures could be very different (Speed, 1993). The males could therefore be honest signallers of unpalatability, and it is conceivable that the mimetic female morphs could still benefit from mimicking more strongly unpalatable species, or that the palatability of the species has decreased recently - after the evolution of the female Batesian mimics. If the andromorphs are recently evolved, it is possible that their evolution is due to this decrease in the palatability of the species, and that they are intraspecific Müllerian mimics. If they are in fact the ancestral form, then their presence in coincidence with the other mimetic morphs would be more difficult to explain, in the absence of other balancing factors.
In order to test the hypothesis that Papilio dardanus is not entirely palatable to predators, experiments were planned in which birds were fed Papilio dardanus, one of its unpalatable models, and palatable butterflies for comparison. In addition, an indirect measure of the palatability of the species was used. It has been shown (Chai & Srygley, 1990; Srygley & Dudley, 1993) that certain morphological traits in butterflies are associated with varying degrees of palatability (as they affect the flight and hence escape capabilities of the insect). Therefore morphological data was collected in accordance with Srygley & Dudley (1993) to assess whether or not Papilio dardanus has the physical structure of an unpalatable butterfly.
The aim of this chapter is to assess the palatability of Papilio dardanus compared with a model for one of the female morphs, Amauris niavius (model for hippocoonides) or Danaus chrysippus (model for lamborni/trophonius). This data should clarify the evolutionary pressures on the species, and be important for the formation of a mathematical model of the populations in Chapter 7.
Hen chicks (Gallus gallus domesticus) have been shown to be discriminating artificial predators, demonstrating aversion to typical unpalatable substances such as quinine and mustard (Gittleman & Harvey, 1980). It was therefore decided to carry out an initial pilot test to determine whether or not they showed aversion to crumbs flavoured with known unpalatable butterflies, or to those flavoured with Papilio dardanus in the same way as their aversion was tested to quinine and mustard by Rowe & Guilford, 1996. If aversion were to be displayed, then further experiments could be carried out to test whether or not they failed to show aversion to known palatable butterflies or whether the aversion was more generally to crumbs flavoured with butterflies.
Dried danaine butterflies have been shown to be highly unpalatable to birds (Dixon et al., 1978), and so refrigerated, desiccated specimens were used to flavour the crumbs.
The aim of this experiment is to determine whether or not hen chicks (Gallus gallus domesticus) can be used to assess the palatability of Papilio dardanus compared with Amauris niavius.
The bodies of two hippocoonides females of Papilio dardanus which had been raised on their natural foodplant (Citrus spp.) in Africa were dessicated over a period of months, and ground into a fine powder. Chick-starter crumbs (Allen & Page, Norfolk Feeds) were damped with a water spray, and then coated in the powder and left under a hot lamp until dry.
The same procedure was carried out with the wings of two hippocoonides females, and the bodies and wings of two Amauris niavius.
This resulted in four different treatments of crumbs:
Three day old hen chicks (Gallus gallus domesticus) from a commercial hatchery which had previously been fed on plain chick-starter crumbs (Allen & Page, Norfolk Feeds), were used in the assessment. The chicks were food deprived for 2-3 hours before the assessments. Two chicks were placed in a circular arena containing a handful of one of the four differently treated groups of crumbs in the centre. Their behaviour was noted, and the time they took to eat the crumbs was recorded. They were then returned to the cage, and their behaviour noted for 30 minutes. This was repeated for each of the four treatments, using different chicks each time.
A few hours later, the chicks were again food-deprived, and put in the arena in pairs. This time they were given the crumbled body and wings of either a desiccated Amauris niavius or a hippocoonides female. Again their behaviour was observed during their time in the arena and for 30 minutes after being returned to the cage. This experiment was repeated twice more the following day with the same chicks receiving the same treatments to ascertain whether or not avoidance had been learnt.
In all four cases, the chicks readily ate all the crumbs presented, and showed no ill effects (such as beak-wiping, head shaking, and large bouts of drinking - Brower et al., 1967). When presented with crumbled butterflies, the chicks ate these just as readily, finishing them within ten minutes. Again, they showed no ill-effects at all. When the experiment was repeated the following day, the chicks appeared to show no aversion to either butterfly, and ate them equally fast with all crumbs being eaten within 10 minutes.
Neither desiccated Amauris niavius nor Papilio dardanus appeared to be distasteful to the young chicks. Therefore they are not suitably discriminative predators to be used to ascertain the relative palatability of Papilio dardanus. Although fresh butterflies may be more unpalatable to the birds, it was not possible to acquire a large number of fresh distasteful butterflies.
Starlings (Sturnus vulgaris) have been used successfully in palatability experiments (Brower, 1960). They were therefore tested in the same way as the hen chicks in Experiment 6-1 to determine whether or not they showed any discrimination between known palatable and unpalatable butterflies.
The aim of this experiment is to determine whether or not starlings (Sturnus vulgaris) can be used to determine the relative palatabilities of Papilio dardanus and one of its models (Amauris niavius).
This experiment used individually caged wild-caught starlings (Sturnus vulgaris) kept in temperature controlled rooms (three cages per room). The starlings were usually fed on turkey crumbs (Allen & Page, Norfolk Feeds). Throughout the experiments the birds were given free access to water.
Initially three starlings were food-deprived for 18 hours and then given 100g turkey crumbs which had been mixed with ground up hippocoonides females of Papilio dardanus (approximately 2 whole butterflies in 100g crumbs) in the method described in Experiment 6-1. Their behaviour was observed for an hour through one-way glass. Secondly, six starlings were food deprived for 17 hours. They were then each given 11g of turkey crumbs - three were plain crumbs and the other three were flavoured with one whole powdered butterfly, one each of Amauris niavius, Papilio dardanus (hippocoonides female), and Aglais urticae (a British nymphalid butterfly, for which there is no evidence of unpalatability). After 6 hours, the amount of food each had eaten and water they had drunk was measured. Finally, two birds were food-deprived for 18 hours and were then given three whole butterflies, roughly crumbled (either Papilio dardanus cenea females or Amauris niavius). Their behaviour was monitored for several hours.
The first three starlings showed no symptoms of distress (such as excessive drinking, beak wiping and head shaking) when fed turkey crumbs mixed with powdered Papilio dardanus. They did show some beak-wiping, and drank regularly, but this was also observed when they were fed plain turkey crumbs.
The birds which were given exactly 11g of treated crumbs ate and drank the following amounts in 6 hours (the amount drunk was assessed by measuring the change in level of the water in the water bottles, in centimeters):
| Starling fed: | Amount eaten | Amount drunk |
|---|---|---|
| plain crumbs | 2.1g | 1.7 units |
| plain crumbs | 7g | 1.7 units |
| plain crumbs | 11g | 5.5 units |
| Amauris niavius crumbs | 0.05g | 1.8 units |
| Papilio dardanus crumbs | 5g | 1.1 units |
| Aglais urticae crumbs | 6.72g | 2 units |
The starling which was given three crumbled Papilio dardanus ate them (including the wings) without signs of particular distress. However, the bird which was given three crumbled Amauris niavius butterflies ate only one piece, then showed prolonged beak-wiping, and did not eat any more of the butterflies.
This small pilot test suggests that the starlings may find turkey crumbs mixed with powdered Amauris niavius unpalatable. It also shows that the starlings will eat 'undisguised' butterflies which have simply been crumbled up, and can thus be induced to take much more concentrated doses of the butterflies (as would a predator in the wild). The reaction of the bird which was given a whole Amauris niavius also suggests that they might find Amauris niavius distasteful. This evidence, although inconclusive, indicates that further experiments should be carried out using starlings as discriminative predators in order to determine whether or not they can be used to place Papilio dardanus on a palatability spectrum ranging from Amauris niavius or Danaus chrysippus to known completely palatable butterflies.
The pilot test, Experiment 6-2 indicated that starlings (Sturnus vulgaris) may find Amauris niavius distasteful. However, it was unfortunately not possible to obtain further supplies of Amauris niavius. Therefore, Danaus chrysippus (the putative model for the trophonius/lamborni morph) was used as an unpalatable butterfly instead. This experiment is designed to test whether or not the starlings can show a great enough degree of discrimination to be able to place Papilio dardanus in the palatability spectrum between Danaus chrysippus, a known unpalatable butterfly, and Charaxes species, shown to be highly palatable by Swynnerton (Carpenter, 1941).
The experiment was designed to test not only their initial reaction to ingesting a particular butterfly, but to then offer the birds a choice to determine whether or not they avoided the species they had just been exposed to, or preferred it to the new species which they had just been introduced to. They were then left with just those two species and no other food, to measure how much of each species they ate as time went on and they became hungrier.
The aim of this experiment is to determine the relative palatabilities of Papilio dardanus, one of its distasteful models, Danaus chrysippus, and palatable Charaxes butterflies using food deprived starlings (Sturnus vulgaris) as discriminative predators.
8 starlings, which had not been used in palatability studies previously, were food deprived for 18 hours, but given open access to water throughout. The starlings were allocated to one of three groups, and each was then given three individual butterflies (which had been refrigerated post mortem for up to 6 months), broken into three pieces and with the wings crumbled into pieces:
- Group A (2 birds) - three Charaxes butterflies each
- Group B (2 birds) - three Danaus chrysippus each
- Group C (4 birds) - three Papilio dardanus females each (one hippocoonides, one lamborni and one cenea for each bird)
The behaviour of each starling was observed for one hour after feeding. At the end of this time, any remaining butterfly material was assessed and the number of pieces eaten by each bird recorded (the crumbled wings were counted as one piece, making the number of pieces of each species 10). The 2 (or 4) birds in each group were then allocated to a sub group, and immediately given a choice of each of two food sources, each prepared in the same way as before:
- Group A1 - choice between Charaxes and Danaus chrysippus
- Group A2 - choice between Charaxes and Papilio dardanus
- Group B1 - choice between Danaus chrysippus and Charaxes
- Group B2 - choice between Danaus chrysippus and Papilio dardanus
- Group C1 - choice between Papilio dardanus and Charaxes
- Group C2 - choice between Papilio dardanus and Danaus chrysippus
The behaviour of each starling was again observed for one hour after feeding, and at the end of this time remaining pieces of butterfly were counted and the amount of each species eaten by each bird recorded as before. The birds were then left with the same choice for 4 further hours, and at the end of this period (a total of 24 hours) any remaining amounts were again counted.
| Bird | First presentation | Second presentation | ||||
|---|---|---|---|---|---|---|
| Species | Amount eaten | Species 1 | Amount eaten | Species 2 | Amount eaten | |
| 1 (A2) | Charaxes | 0 | Charaxes | 0 | Papilio | 0 |
| 2 (A1) | Charaxes | 8 | Charaxes | 10 | Danaus | 10 |
| 3 (C1) | Papilio | 10 | Papilio | 10 | Danaus | 6 |
| 4 (C2) | Papilio | 10 | Papilio | 5 | Charaxes | 7 |
| 5 (C1) | Papilio | 2 | Papilio | 10 | Danaus | 10 |
| 6 (C2) | Papilio | 0 | Papilio | 10 | Charaxes | 10 |
| 7 (B2) | Danaus | 0 | Danaus | 10 | Papilio | 10 |
| 8 (B1) | Danaus | 0 | Danaus | 10 | Charaxes | 10 |
The starlings appeared to be indiscriminate in what they ate, most eating everything they were presented with. This could either be because the dried specimens did not differ in palatability to the birds, or because the hunger levels of the birds were so high as to cause them not to distinguish between the palatabilities of what they were presented with. As mentioned previously, large numbers of fresh specimens of the butterflies (other than Papilio dardanus) were very difficult to obtain, and so the experiment was repeated with a shorter food deprivation time in order to decrease the hunger levels of the birds.
In Experiment 6-3, the starlings failed to discriminate between known palatable (Charaxes) and known unpalatable (Danaus chrysippus) butterflies. This could be due to the dried butterfly specimens losing their unpalatability. However, it was not possible to obtain a large number of fresh specimens of a known unpalatable species. An alternative is that the hunger levels of the starlings were too high to allow them to discriminate between the species. Therefore, another test was carried out, repeating the general method of Experiment 6-4 but decreasing the food deprivation period of the experimental birds.
The aim of this experiment is to determine whether or not starlings with a lower level of hunger will discriminate between known palatable (Charaxes) and unpalatable (Danaus chrysippus) butterflies, and also to test the relative palatability of Papilio dardanus with respect to these two extremes.
The same basic method of Experiment 6-3 was used. 6 starlings, not previously used in palatability experiments, were food deprived for 5 hours, but given free access to water during this period. The starlings were then allocated to one of three groups, and each was then given three individual butterflies (which had been refrigerated post mortem for up to one year), each broken into three pieces and with the wings crumbled into pieces:
- Group A - three Charaxes butterflies each
- Group B - three Danaus chrysippus each
- Group C - three Papilio dardanus females each (one hippocoonides, one lamborni and one cenea)
The behaviour of each starling was observed for one hour after feeding. At the end of this time, any remaining butterfly pieces were counted and the amount eaten by each bird recorded (the crumbled wings were counted as one piece, giving each bird a score out of 10 for each butterfly species). The 2 birds in each group were then allocated to a sub group, and given a choice of each of two butterflies, prepared in the same way as in the first part of the experiment:
- Group A1 - choice between Charaxes and Danaus chrysippus
- Group A2 - choice between Charaxes and Papilio dardanus
- Group B1 - choice between Danaus chrysippus and Charaxes
- Group B2 - choice between Danaus chrysippus and Papilio dardanus
- Group C1 - choice between Papilio dardanus and Charaxes
- Group C2 - choice between Papilio dardanus and Danaus chrysippus
The behaviour of each starling was again observed for one hour after feeding, and at the end of this time the number of pieces of each butterfly remaining and the amount of each species eaten by each bird were recorded as before. The birds were then left with the same choice for 4 further hours, and at the end of this period (a total of 11 hours) any remaining pieces were again counted.
| Bird | First presentation | Second presentation | ||||
|---|---|---|---|---|---|---|
| Species | Amount eaten | Species 1 | Amount eaten | Species 2 | Amount eaten | |
| 1 (A1) | Charaxes | 0 | Charaxes | 0 | Danaus | 0 |
| 2 (A2) | Charaxes | 9 | Charaxes | 10 | Papilio | 10 |
| 3 (C2) | Papilio | 9 | Papilio | 0 | Danaus | 3 |
| 4 (C1) | Papilio | 6 | Papilio | 1 | Charaxes | 3 |
| 5 (B2) | Danaus | 10 | Danaus | 10 | Papilio | 10 |
| 6 (B1) | Danaus | 10 | Danaus | 10 | Charaxes | 9 |
As in Experiment 6-3, there appears to be no pattern to the feeding of the starlings. Some ate almost all that was presented to them, and others ate very little. The birds did not appear to avoid the unpalatable Danaus chrysippus. The observed difference in behaviour could be due to differing hunger levels in the individuals. With relatively short periods of food deprivation it is very difficult to ensure that all the birds are in the same state. However, even those that did not eat everything presented to them appeared not to avoid the Danaus chrysippus (e.g. bird 3) and hence it appears that the dried butterfly specimens have lost any unpalatability they may have had for the birds, or that the birds do not find the chemicals contained in Danaus chrysippus (thought to be cardenolides and pyrrolizidine alkaloids - Meinwald et al., 1974) distasteful. Swynnerton (1915, 1919) found some birds very sensitive to the compounds in Danaus chrysippus (one wood hoopoe, Phoeniculus purpureus, vomiting after eating a single Danaus chrysippus, and another after eating a single Amauris echeria - the model for the cenea morph), whilst others seemed completely insensitive (rollers, Coracias garrulus; yellow-billed hornbills, Tockus flavirostrus; crowned hornbills, Tockus alboterminatus; and babblers, Turdoides jardinei ate up to 50 danaines without vomitting). This could be due to cardenolide insensitivity (Brower, 1984) or due to the presence of cardenolide-free danaines in the population (Brower et al., 1975, 1978). The same explanations could be true for the starlings not finding the danaines unpalatable.
Experiment 6-4 indicated that starlings (Sturnus vulgaris) did not find dried specimens of either Papilio dardanus or Danaus chrysippus unpalatable, but it was not possible to determine whether this was due to the dried nature of the butterflies, whether the birds were resistant to the effects of the cardenolides and pyrrolizidine alkaloids of Danaus chrysippus (Brower, 1984), or whether the individual butterflies were lacking in protection (Brower et al., 1975, 1978). It was possible to obtain a small number of fresh Danaus chrysippus from Malaysia, and so it was possible to design a small experiment to test which of these factors was causing the starlings to be indiscriminate.
The aim of this experiment is to determine whether or not starlings (Sturnus vulgaris) find fresh Danaus chrysippus or Papilio dardanus distasteful.
Experiment 6-4 illustrated how a short food deprivation period could result in different levels of hunger in different individuals, leading to very different degrees of discrimination. A small pilot test indicated that the starlings would eat small pieces of butterfly even without a period of food deprivation, and so the experiment was designed without deprivation, giving the starlings a choice between their normal dried turkey crumbs (Allen & Page, Norfolk Feeds) and small pieces of butterfly.
Six starlings (the same birds as used in Experiment 6-4), fed normally on turkey crumbs (Allen & Page, Norfolk Feeds) and given free access to water at all times were allocated randomly to one of two groups. The three birds in Group A were each given one freshly killed Papilio dardanus, the body divided into five pieces, in one half of a dual feeder, and a supply of their normal turkey crumbs in the other half. The birds in Group B were given one freshly killed Danaus chrysippus, divided in the same manner, in one half of the feeder, and a supply of turkey crumbs in the other half.
Both sets of birds were left for four hours, after which time the number of pieces of butterfly they had eaten was counted (recorded as a score out of 5). Any remaining pieces were then removed, and the birds left with just their normal food.
This procedure was then repeated twice more, at 24 hour intervals, with the groups remaining the same as on the previous day, and the number of pieces of the butterflies eaten by each starling on each day recorded.
The amounts eaten by each starling on each day are shown in Table 6-4.
| Bird number | Number of pieces eaten | |||
|---|---|---|---|---|
| Day 1 | Day 2 | Day 3 | ||
| Group A (Papilio dardanus) | 1 | 5 | 5 | 5 |
| 2 | 5 | 5 | 5 | |
| 3 | 5 | 5 | 5 | |
| Group B (Danaus chrysippus) | 4 | 1 | 0 | 0 |
| 5 | 5 | 5 | 5 | |
| 6 | 1 | 5 | 5 | |
The small numbers of birds and butterflies used makes the results difficult to interpret. However, the results suggest that the fresh Danaus chrysippus may not be quite as palatable as the fresh Papilio dardanus to the starlings although bird 4 in this experiment, which ate very little of the Danaus chrysippus is the same bird as Bird 1 in Experiment 6-4, which ate none of the butterflies presented (although it did eat mealworms presented in the same food hopper between experiments). The Danaus chrysippus did not have any emetic effect on the starlings, and the birds did not appear to find them particularly distasteful (as they ate the butterflies without any food deprivation, and whilst there was other food available). It thus appears that either the starlings are not affected by the distasteful compounds in Danaus chrysippus or these individual butterflies come from a population which does not contain such compounds. It appears clear, however, that Papilio dardanus is highly palatable to the starlings (it has been eaten in every experiment, and in this experiment it was eaten in preference to the turkey crumbs).
It has been shown that some morphological traits, such as thoracic diameter, abdominal length, and the position of the centre of mass of the body can be used as predictors of palatability (Chai & Srygley, 1990; Srygley & Dudley, 1993, Srygley, 1994). This is due to the fact that palatable butterflies tend to allocate more resources to flight muscle in the thorax (increasing the thoracic diameter) enhancing flight speed and acceleration in case of attack by birds (Chai & Srygley, 1990). They also have shorter abdomens, possibly in order to decrease the chance of being caught by the abdomen and to increase aerobatic ability (Chai & Srygley, 1990). Furthermore, they tend to have a centre of mass positioned closer to the wing base than unpalatable butterflies, which increases the manoeuvrability of the butterflies, allowing them to escape from predators (Srygley & Dudley, 1993). The slender abdominal profile of unpalatable butterflies may additionally be used as a signal of distastefulness (Chai & Srygley, 1990), and therefore some Batesian mimics may attempt to simulate this (Srygley, 1994).
The aim of this experiment is to determine whether or not the hippocoonides morphs or the males are likely to be palatable or unpalatable on the basis of their morphology.
Ten male and nine hippocoonides female Papilio dardanus were used, and these were compared with ten Danaus chrysippus, following the general method of Chai & Srygley (1990) followed. Measurements of the full body length of each specimen were made using a pair of Manostat measuring callipers (accurate to 0.05mm) by holding the abdomen of the live butterflies out straight, in line with the thorax, and measuring the length from the front of the head (not measuring the antennae) to the end of the genitalia. The width of the thorax was then measured by holding the butterflies' wings together dorsally, and tightening the callipers until they gripped the sides of the thorax at the point where the forewings joined the body. The ratio of body length to thorax width was then calculated for each specimen.
To ascertain the centre of mass for males and six of the hippocoonides females (three did not emerge properly from their pupal cases, and their abdomens were full of fluid), the bodies of freshly killed specimens were first weighed on an Oertling NA264 balance (accurate to 0.1mg). Each body was then divided into several parts. First the head length was measured. This was done most accurately by measuring the diameter of the eye, which comprises most of the head. The head was then removed and weighed. The length of the thorax was measured next. The wings were then removed, the abdomen snipped off, and the thorax weighed. Finally, the abdomen was divided into three parts - the first, approximately three segments and containing the crop, the second approximately two segments containing the reproductive tracts, and the final part comprising three segments and containing the sclerotised genetalia. Each length of abdomen was measured, and then weighed.
To calculate the centre of mass of the insect, each section which had been weighed and measured was assumed to be of uniform density, and that the centre of mass lay on the axis of symmetry of the body. This assumption meant that the centre of mass of each section lay halfway down its length, and hence the turning force of each section around the head end could be calculated. The distance of the centre of mass of the entire insect from the head was thus the sum of these turning moments divided by the total mass of the insect (see Figure 6-2).
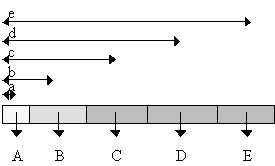
Centre of mass of entire insect is situated a distance m from the head where:
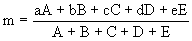
If the wing base of the insect is assumed to be in the centre of the thorax (distance b from the head), then the centre of mass of the insect is situated a distance (m - b) from the wing base. This distance can then be expressed as a fraction of the total length of the body.
Thorax width: body length ratio.
| Male Specimen | Thorax width (mm) | Body length (mm) | Ratio |
|---|---|---|---|
| 1 | 4.05 | 27.05 | 6.679 |
| 2 | 3.60 | 24.25 | 6.736 |
| 3 | 4.00 | 24.55 | 6.063 |
| 4 | 4.10 | 28.80 | 7.024 |
| 5 | 3.85 | 25.55 | 6.636 |
| 6 | 3.35 | 24.00 | 7.164 |
| 7 | 4.65 | 28.20 | 6.065 |
| 8 | 4.75 | 27.55 | 5.800 |
| 9 | 5.10 | 26.00 | 5.098 |
| 10 | 4.45 | 27.95 | 6.281 |
| MEAN | 4.19 | 26.36 | 6.355 |
| Female Specimen | Thorax width (mm) | Body length (mm) | Ratio |
|---|---|---|---|
| 1 | 5.10 | 29.05 | 5.696 |
| 2 | 4.80 | 27.05 | 5.635 |
| 3 | 4.85 | 27.10 | 5.588 |
| 4 | 3.70 | 26.10 | 7.054 |
| 5 | 4.35 | 31.45 | 7.230 |
| 6 | 4.65 | 39.80 | 8.559 |
| 7 | 4.85 | 26.10 | 5.381 |
| 8 | 4.20 | 25.15 | 5.988 |
| 9 | 4.40 | 24.20 | 5.500 |
| MEAN | 4.54 | 28.44 | 6.292 |
| Specimen | Sex | Thorax width (mm) | Body length (mm) | Ratio |
|---|---|---|---|---|
| 1 | F | 4.15 | 23.40 | 5.639 |
| 2 | F | 3.95 | 22.35 | 5.658 |
| 3 | M | 4.1 | 23.40 | 5.707 |
| 4 | F | 3.85 | 23.60 | 6.130 |
| 5 | M | 4.2 | 24.20 | 5.762 |
| 6 | M | 3.8 | 25.05 | 6.592 |
| 7 | F | 3.7 | 22.1 | 5.973 |
| 8 | M | 3.95 | 25.15 | 6.367 |
| 9 | M | 4.15 | 23.55 | 5.675 |
| 10 | M | 3.7 | 22.2 | 6.000 |
| MEAN | 3.96 | 23.50 | 5.923 |
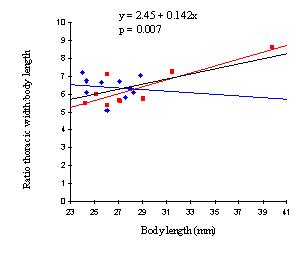
A graph of the ratio of thoracic width to body length for males and females of Papilio dardanus shows that the ratio does increase slightly with body length (p=0.007, see Figure 6-3), but there is no significant difference between the sexes (p=0.487, Mann-Whitney test, two-tailed).
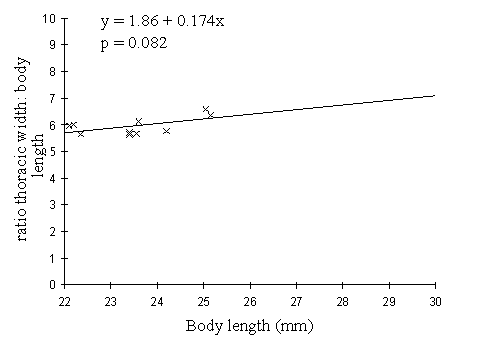
For Danaus chrysippus, there is a similar increase in ratio with body length, although this is not significant (p=0.082), and the body length shows much less variation. There is no significant difference between the sexes (p=0.337, Mann-Whitney test, two-tailed).
Chai & Srygley (1990) plot body shape (the ratio of body length to thoracic width) against palatability (as measured by percentage eaten in feeding experiments) for 53 species of butterfly. This graph shows that highly palatable butterflies tend to have a ratio of between 4 and 6, and unpalatable butterflies tend to have ratios of 8.5 and above (with a couple of exceptions). At intermediate values, where both Papilio dardanus and Danaus chrysippus appear to lie, the butterflies tend to have palatabilities of below 50%. Figure 6-3 however, indicates that body length must be taken into account, as, at least in some species, butterflies with longer bodies tend to have a higher ratio. Therefore comparing two species with differing overall body length may give misleading results. In this case, Danaus chrysippus is a smaller butterfly, and thus the ratio of body length to thoracic width may be lower than expected for an unpalatable species.
| Turning force (mgmm) around head of centre of mass of: | distance centre mass from: | |||||||
|---|---|---|---|---|---|---|---|---|
| Head | Thorax | Abdomen 1 | Abdomen 2 | Abdomen 3 | Head | Wing base | ||
| Male | 1 | 18.59 | 393.36 | 141.44 | 403.66 | 423.15 | 0.411 | 0.167 |
| 2 | 14.5 | 259.55 | 86.45 | 305.61 | 277.38 | 0.417 | 0.168 | |
| 3 | 23.40 | 484.22 | 202.06 | 614.56 | 473.67 | 0.429 | 0.173 | |
| 4 | 16.06 | 544.36 | 243.89 | 962.32 | 279.38 | 0.379 | 0.164 | |
| 5 | 13.51 | 363.20 | 141.74 | 478.98 | 373.36 | 0.394 | 0.172 | |
| 6 | 8.19 | 274.80 | 68.15 | 280.79 | 274.3 | 0.411 | 0.172 | |
| 7 | 20.13 | 590.29 | 186.73 | 928.25 | 393.53 | 0.409 | 0.174 | |
| 8 | 23.03 | 580.97 | 490.78 | 827.52 | 528.41 | 0.442 | 0.187 | |
| 9 | 14.15 | 405.38 | 231.42 | 398.36 | 1045.23 | 0.402 | 0.130 | |
| 10 | 21.17 | 697.50 | 293.15 | 605.01 | 864.97 | 0.333 | 0.083 | |
| MEAN | 17.27 | 459.36 | 208.58 | 580.51 | 493.34 | 0.403 | 0.159 | |
| Female | 1 | 23.00 | 680.21 | 716.94 | 1343.65 | 1293.86 | 0.478 | 0.245 |
| 2 | 15.93 | 578.10 | 585.09 | 1113.81 | 971.19 | 0.525 | 0.264 | |
| 3 | 16.25 | 509.72 | 308.36 | 1058.64 | 818.35 | 0.521 | 0.260 | |
| 4 | 10.35 | 401.79 | 417.15 | 645.44 | 713.45 | 0.458 | 0.232 | |
| 5 | 15.89 | 857.21 | 1156.32 | 2155.43 | 2169.14 | 0.486 | 0.281 | |
| 6 | 9.90 | 539.28 | 810.65 | 2049.27 | 1127.30 | 0.386 | 0.228 | |
| MEAN | 15.22 | 594.38 | 665.75 | 1394.37 | 1182.21 | 0.476 | 0.252 | |
| Specimen | Sex | Turning force (mgmm) around head of centre of mass of: | distance centre mass from: | |||||
|---|---|---|---|---|---|---|---|---|
| Head | Thorax | Abdomen 1 | Abdomen 2 | Abdomen 3 | Head | Wing base | ||
| 1 | F | 9.0 | 461.76 | 418.5 | 934.95 | 174.3 | 0.432 | 0.210 |
| 2 | F | 6.6 | 289.44 | 226.4 | 493.65 | 154.8 | 0.403 | 0.389 |
| 3 | M | 8.4 | 445.72 | 203.4 | 487.5 | 359.9 | 0.389 | 0.174 |
| 4 | F | 7.91 | 357.64 | 233.2 | 680.52 | 207.6 | 0.387 | 0.189 |
| 5 | M | 8.50 | 495.33 | 255.0 | 433.92 | 402.6 | 0.406 | 0.170 |
| 6 | M | 7.65 | 516.88 | 308.73 | 716 | 322.64 | 0.414 | 0.190 |
| 7 | F | 7.28 | 293.76 | 172.37 | 422.30 | 273.75 | 0.408 | 0.190 |
| 8 | M | 9.18 | 608.06 | 363.05 | 719.82 | 475.58 | 0.437 | 0.203 |
| 9 | M | 11.99 | 563.55 | 326.12 | 689.27 | 341.94 | 0.414 | 0.179 |
| MEAN | 8.50 | 448.02 | 278.55 | 619.77 | 301.46 | 0.410 | 0.188 | |
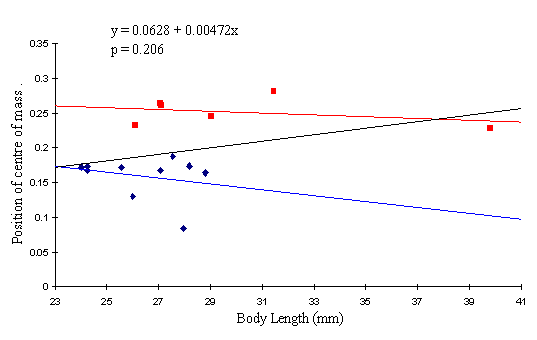
A graph of the data for Papilio dardanus (see Figure 6-5) shows that the position of the centre of mass for the species does not vary significantly with the length of the body (p=0.206), and shows that females have a centre of mass set consistently further posterior than males (p=0.0014; Mann-Whitney test, two-tailed).
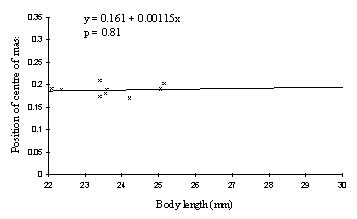
A graph of the data for Danaus chrysippus (see Figure 6-6) indicates that again the centre of mass does not vary significantly with body length (p=0.81), and also that the position of the centre of mass in females is not significantly different from that in males (p=0.27; Mann-Whitney test, two-tailed). The females of Papilio dardanus, however, have a centre of mass positioned significantly further back than Danaus chrysippus (p=0.0018; Mann-Whitney test, two-tailed).
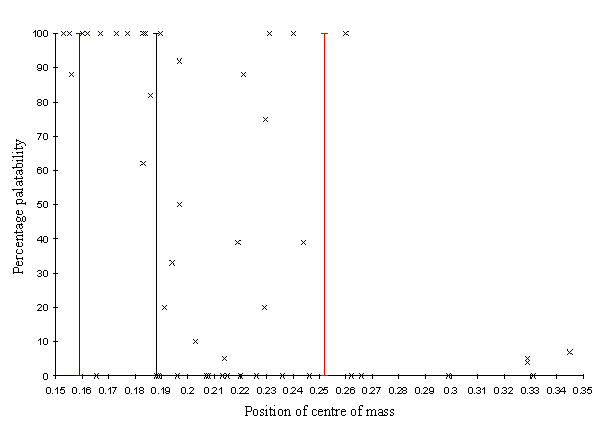
Srygley and Dudley (1992), and Srygley (1994) give data for the percentage palatability and the position of the centre of mass for 51 species of butterfly. A graph of this can be seen in Figure 6-7. The highly palatable butterflies tend to cluster with a centre of body mass between 0.15 and 0.19 body-length units from the wing base. The mid- to unpalatable species tend to have centres of mass between 0.19 and 0.25 (the Batesian mimics are reported to be at the extreme end of this distribution, with centres of mass between 0.2-0.25 body-units from the wing base). There is another cluster of unpalatable butterflies with a centre of body mass greater than 0.3 body lengths from the wing base.
The overall species mean position of the centre of mass (0.194 body lengths from the wing base) for Papilio dardanus therefore falls within the distribution of the mid- to unpalatable butterflies. The males are in the range for highly palatable butterflies and the females being in the normal range for palatable, Batesian mimics. The mean position of the centre of mass in Danaus chrysippus also falls in the range for mid- to unpalatable butterflies.
The results of the both sets of data gathered for Papilio dardanus indicates that it is a Batesian mimic. From the evidence of the position of the centre of mass, it appears that the females of Papilio dardanus may not have shifted their centre of mass slightly in order to mimic their unpalatable models (Srygley, 1994), since their centre of mass is actually positioned further back than that of one of their models, Danaus chrysippus, hence putting them at greater risk than would be necessary to mimic the shape of their models. This effect is likely to be due simply to the heavy load of eggs carried by the females. Srygley (1994) did not analyse data for males and females separately, and so in order to compare the data for Papilio dardanus to that presented in that paper (Srygley, 1994) it is necessary to take a mean value for the species.
The fact that the centre of mass is set further posterior in female Papilio dardanus means that they will not be as aerobatically manoeuvrable as males (Srygley & Dudley, 1993). This disadvantage could be a contributory factor causing female butterflies to be mimetic in species where males are not. Not only is there an evolutionary pressure for them to survive longer and lay more eggs (whilst males only need to mate), but the egg load they carry, by moving the centre of mass away from the wing base, causes them to be more prone to attack from birds (Srygley & Dudley, 1993). This is in accordance with data on wing damage collected by Ohsaki (1995), who found that non-mimetic females of Papilio polytes appeared to suffer more bird attacks than males (although mimetic females appeared to receive as few attacks as males and their models, Pachiliopta aristolochiae). The fact that females of Danaus chrysippus did not have a significantly different positioning of the centre of mass is interesting. It is possible that different egg loads carried by females of different species can create different evolutionary pressures on the sexes in some species, but not in others.
Papilio dardanus appears to be completely palatable to starlings. Danaus chrysippus, however, also appeared to be palatable to the starlings, although it is possible that the individual butterflies used were from a population which did not sequester cardiac glycosides efficiently. The measurements of the morphology of the two species indicated that they both lie in a region where it is difficult to predict the palatability with any accuracy. However, the males of Papilio dardanus had a centre of mass which indicated that they were completely palatable, and therefore it seems likely that the species as a whole is palatable.
The morphology of the butterflies also demonstrated that the females of Papilio dardanus are likely to be less manoeuvrable than the males in flight, due to the position of their centre of mass. This is likely to make them much easier prey for birds to catch (Srygley & Dudley, 1993), and hence increases the pressure on females to avoid attack. This could be a contributing factor to the phenomenon of female-limited mimicry in butterflies. Turner (1978) reviewed this topic of debate, and concluded that there was no evidence that female butterflies were subject to more predation than males, but the evidence from Ohsaki (1995), and the sex difference in the degree of flight manoeuvrability now suggests that they are. The theory that female choice maintains male polymorphism (Belt, 1874), which was favoured by Turner (1978) has very little evidence to support it (female choice in butterflies appears only to have been demonstrated in Danaus chrysippus, in which both sexes are in fact polymorphic; Smith, 1984). In Papilio dardanus, the female-limited mimicry is spectacular. Male sexual selection appears to be operating (Cook et al., 1994 and see also Chapter 5), whilst female choice has not yet been demonstrated (see Chapter 5). The females fly under thicker cover (Nigel Venters , pers. comm.), and at different times of day from the males (Cook et al., 1994), and the measurements taken in Experiment 6-6 indicate that they are likely to be more easily caught by birds. The evidence, therefore, for Papilio dardanus, suggests that an increased predation risk for females may be a more important factor in the evolution of female-limited mimicry than the action of female choice on male coloration. The evidence of the position of the centre of mass in Danaus chrysippus indicates that in this species there is no difference between the aerobatic ability of the males and the females. Therefore in this species the evolutionary pressures on the two sexes are likely to be much more similar. Differences in the egg laying strategies of different species may therefore have a profound effect on the evolutionary pressures on the two sexes, and may be an explanation for the existence of sex-limited mimicry in some species and its absence in others. Measurements by Srygley (1994) from mimetic and non-mimetic species showed that Batesian mimics on the whole tended to have a centre of mass positioned slightly further back than non-mimics. He interpreted this as possible evidence of 'locomotor mimicry'. However, it is also possible that the position of the centre of mass of those species had meant that they were more likely to evolve mimicry, due to the fact that they were slightly less agile in flight. Further measurements of both sexes in a range of sex-limited and sex-unlimited mimetic species and their models may help clarify this issue.
Many thanks to everyone who helped supply butterflies for this experiments, especially John Calvert at Stratford Butterfly Farm and Pierre Le Roux in South Africa. Many thanks also to Dave Wilson for providing and looking after the starlings, and to Julian Howe for the chicks.
I am also very grateful to Bob Srygley for suggesting the method for taking measurements of the butterflies, and for the loan of the calipers, and to Nicky Warren for the use of the balance.
Belt, T. 1874. The naturalist in Nicaragua. Murray, London.
Bernardi, G. 1963. Quelques aspects zoogéographiques du mimétisme chez les Lépidpidoptčres. Proceedings, 16th International Congress of Zoology, Washington 4, 161-166.
Brower, J.V.Z. 1960. Experimental studies of mimicry. IV The reactions of starlings to different proportions of models and mimics. Am. Nat. 94, 271-282.
Brower, L.P. 1969. Ecological chemistry. Scientific American, 22-29.
Brower, L.P. 1984. Chemical defence in butterflies. In The Biology of Butterflies (ed. Vane-Wright, R.I. & Ackery, P.R.) Princetown University Press, New Jersey.109-134.
Brower, L.P., Brower, J.V.Z. & Corvino, J.M. 1967. Plant poisons in a terrestrial food chain. Proc. Natn. Acad. Sci. U.S.A. 57, 893-898.
Brower, L.P., Edmunds, M. & Moffitt, C.M. 1975. Cardenolide content and palatability of a population of Danaus chrysippus butterflies from West Africa. J. Ent. (A) 49, 183-196.
Brower, L.P., Gibson, D.O., Moffitt, C.M. & Panchen, A.L. 1978. Cardenolide content of Danaus chrysippus butterflies from three areas of East Africa. Biol. J. Linn. Soc. 10, 251-273.
Brower, L.P., Ryerson, W.N., Coppinger, L.L., & Glazier, S.C. 1968. Ecological chemistry and the palatability spectrum. Science 161, 1349-51.
Carpenter, G.D.H. 1941. Observations and experiments in Africa by the late C.F.M. Swynnerton on wild birds eating butterflies and the preference shown. Proc. Linn. Soc. Lond. 154, 10-46.
Chai, P. & Srygley, R.B. 1990. Predation and the flight, morphology and temperature of Neotropical rain-forest butterflies. Am. Nat. 135, 748-765.
Charlesworth, D. & Charlesworth, B. 1976. Theoretical genetics of Batesian mimicry III Evolution of dominance. J. Theor. Biol. 55, 325-337.
Clarke, C.A., Clarke, F.M.M., Collins, S.C., Gill, A.C.L., Turner, J.R.G. 1985. Male-like females, mimicry, and transvestism in swallowtail butterflies. Systematic Entomology 10, 257-283.
Clarke, C.A. & Sheppard, P.M. 1963. Interactions between major genes and polygenes in the determination of the mimetic patterns of Papilio dardanus. Evolution 17, 404-413.
Cook, S.E., Vernon, J.G., Bateson, M., Guilford, T. 1994. Mate choice in the polymorphic African swallowtail butterfly, Papilio dardanus: male-like females may avoid sexual harrassment. Animal Behaviour 47, 389-397.
Dixon, C.A., Erikson, J.M., Kellett, D.N. & Rothschild, M. 1978. Some adaptations between Danaus plexippus and its food plant, with notes on Danaus chrysippus and Euploea core. J. Zool., Lond. 185, 437-467.
Fink, L.S. & Brower, L.P. 1981. Birds can overcome the cardenolide defences of Monarch butterflies in Mexico. Nature 291, 67.
Ford, E.B. 1936. The genetics of Papilio dardanus Brown (Lep.). Transactions of the Royal Entomological Society of London 85, 435-466.
Ford, E.B. 1955. Moths. London.
Gittleman, J.L. & Harvey, P.H. 1980. Why are distasteful prey not cryptic? Nature 286, 149-150.
Glendinning, J.I. & Brower, L.P. 1990. Feeding and breeding responses of five mice species to overwintering aggregations of the Monarch butterfly. Journal of Animal Ecology 59, 1091-1112.
Meinwald, J., Boriack, C.J., Schneider, D., Boppré, M., Wood, W.F. & Eisner, T. 1974. Volatile ketones in the hairpencil secretion of danaid butterflies (Amauris & Danaus), Experientia 30, 721-722.
O'Donald, P. & Barrett, J.A. 1973. Evolution of dominance in polymorphic Batesian mimicry. Theoretical Population Biology 4, 173-192.
Ohsaki, N. 1995. Preferential predation of female butterflies. Nature 378, 173-5.
Poulton, E.B. 1924. Papilio dardanus. The most interesting butterfly in the world. Journal of the East African and Ugandan Natural History Society 20, 4-22.
Rothschild, M., Moore, B.P. & Brown, W.V. 1984. Pyrazines as warning odour components in the monarch butterfly, Danaus plexippus, and in moths of the genera Zygaena and Amata. Biol. J. Linn. Soc. 23, 375-380.
Rowe, C. & Guilford, T. 1996. Hidden colour aversions in domestic chicks triggered by pyrazine odours of insect warning displays. Nature 383, 520-522.
Sheppard, P.M. 1967. Natural selection and heredity (3rd edn.). Hutchinson, London.
Speed, M.P. 1993. Müllerian mimicry and the psychology of predation. Anim. Behav. 45, 571-580.
Srygley, R.B. 1994. Locomotor mimicry in butterflies? The associations of positions of centres of mass among groups of mimetic, profitable prey. Phil. Trans. R. Soc. Lond. B 343, 145-155.
Srygley, R.B. & Dudley, R. 1993. Correlations of the position of center of body mass with butterfly escape tactics. J. Exp. Biol. 174, 155-166.
Srygley, R.B. & Kingsolver, J.G. 1998. Red-wing blackbird reproductive behaviour and the palatability, flight performance, and morphology of temperate pierid butterflies (Colias, Pieris, and Pontia). Biol. J. Linn. Soc. 64 (1), 41-55.
Swynnerton, C.F.M. 1915. Birds in relation to their prey. Experiments on wood-hoopoes, small hornbillsa, and a babbler. Jl. S. Afr. Orn. Un. 1915, 22-108.
Swynnerton, C.F.M. 1919. Experiments and observations bearing on the explanation of form and colouring, 1908-1913, Africa. Zool. J. Linn. Soc. 33, 203-385.
Trimen, R. 1868. On some remarkable mimetic analogies among African butterflies. Transactions of the Linnean Society of London 26, 497-522.
Turner, J.R.G. 1963. Geographical variation and evolution in the males of the butterfly Papilio dardanus Brown (Lepidoptera: Papilionidae). Trans. Roy. Ent. Soc. Lond. 115, 239-259.
Turner, J.R.G. 1978. Why male butterflies are non-mimetic: natural selection, sexual selection, group selection, modification and sieving. Biol. J. Linn. Soc. 10, 385-432.
Van Bemmelen, J.F. 1922. The wing-design of mimetic butterflies. Proceedings, Section of Sciences, Koninklijke Nederlandse Akademie Wetensschappen 23, 877-886.
Vane-Wright, R.I., Raheem, D.C., Cieslak, A., Vogler, A.P. In press. Evolution of the mimetic African swallowtail butterfly Papilio dardanus: molecular data confirms relationships with P. phorcas and P. constantinus. Biol. J. Linn. Soc.
Vane-Wright, R.I. & Smith, C.R. 1991. Phylogenetic relationships of three African swallowtail butterflies, Papilio dardanus, P. phorcas, and P. constantinus: a cladistic analysis (Lepidoptera: Papilionidae). Systematic Entomology 16: 275-291.
|
Please cite this thesis as: Freeman, ALJ; 1998; D.Phil thesis, Oxford University. |
E-mail to Alexandra Freeman Back to Table of Contents Back to Home Page |